วิวัฒนาการของมนุษย์
วิวัฒนาการของมนุษย์ (อังกฤษ: Human evolution) เป็นกระบวนการวิวัฒนาการที่นำไปสู่การปรากฏขึ้นของ "มนุษย์ปัจจุบัน" (อังกฤษ: modern human มีนามตามอนุกรมวิธานว่า Homo sapiens หรือ Homo sapiens sapiens) ซึ่งแม้ว่าจริง ๆ แล้วจะเริ่มต้นตั้งแต่บรรพบุรุษแรกของสิ่งมีชีวิตทั้งหมด แต่บทความนี้ครอบคลุมเพียงแค่ประวัติวิวัฒนาการของสัตว์อันดับวานร (primate) โดยเฉพาะของสกุล โฮโม (Homo) และการปรากฏขึ้นของมนุษย์สปีชีส์ Homo sapiens ที่จัดเป็นสัตว์วงศ์ลิงใหญ่เท่านั้น การศึกษาเกี่ยวกับวิวัฒนาการมนุษย์นั้นต้องอาศัยความรู้ทางวิทยาศาสตร์หลายสาขา รวมทั้งมานุษยวิทยาเชิงกายภาพ (หรือ มานุษยวิทยาเชิงชีวภาพ), วานรวิทยา, โบราณคดี, บรรพชีวินวิทยา, พฤติกรรมวิทยา, ภาษาศาสตร์, จิตวิทยาเชิงวิวัฒนาการ (evolutionary psychology), คัพภวิทยา และพันธุศาสตร์[1]

กระบวนการวิวัฒนาการเป็นความเปลี่ยนแปลงของลักษณะสืบสายพันธุ์ (trait) ของกลุ่มสิ่งมีชีวิตผ่านหลายชั่วยุคชีวิต เป็นกระบวนการที่ทำให้เกิดความหลายหลากกับสิ่งมีชีวิตในทุกระดับชั้น รวมทั้งระดับสปีชีส์ ระดับสิ่งมีชีวิตแต่ละชีวิต และแม้กระทั่งโครงสร้างระดับโมเลกุลเช่นดีเอ็นเอและโปรตีน[2] สิ่งมีชีวิตทั้งหมดในโลกสืบสายมาจากบรรพบุรุษเดียวกันที่มีชีวิตประมาณ 3.8 พันล้านปีก่อน การเกิดสปีชีส์ใหม่ ๆ และการแยกสายพันธุ์ออกจากกันของสิ่งมีชีวิต สามารถอนุมานได้จากลักษณะสืบสายพันธุ์ทางสัณฐานและทางเคมีชีวภาพ หรือโดยลำดับดีเอ็นเอที่มีร่วมกัน[3] คือ ลักษณะสืบสายพันธุ์และลำดับดีเอ็นเอที่มีกำเนิดเดียวกัน จะมีความคล้ายคลึงกันระหว่างสปีชีส์ที่มีบรรพบุรุษร่วมกันเร็ว ๆ นี้มากกว่าระหว่างสปีชีส์ที่มีบรรพบุรุษร่วมกันมานานแล้ว ดังนั้นความคล้ายคลึงกันและความแตกต่างกันจึงสามารถใช้สร้างแบบของต้นไม้สายพันธุ์สิ่งมีชีวิต ที่แสดงความสัมพันธ์เชิงญาติ โดยใช้สิ่งมีชีวิตที่ยังมีอยู่หรือใช้ซากดึกดำบรรพ์เป็นหลักฐานข้อมูล รูปแบบความหลากหลายของสิ่งมีชีวิตในโลกเปลี่ยนแปลงไปเพราะการเกิดขึ้นของสปีชีส์ใหม่ ๆ และการสูญพันธุ์ไปของสิ่งมีชีวิตที่มีอยู่[4]
งานวิจัยต่าง ๆ ทางพันธุศาสตร์แสดงว่า สัตว์อันดับวานรรวมทั้งมนุษย์แยกออกจากสัตว์เลี้ยงลูกด้วยนมประเภทอื่น ๆ เมื่อประมาณ 85 ล้านปีก่อน โดยมีซากดึกดำบรรพ์ปรากฏเป็นครั้งแรกสุดเมื่อประมาณ 55 ล้านปีก่อน[5] ส่วนลิงวงศ์ชะนี (Hylobatidae) แยกสายพันธุ์ออกจากสายพันธุ์วงศ์ลิงใหญ่ (Hominidae) รวมทั้งมนุษย์ ซึ่งเป็นวงศ์หนึ่ง ๆ ของสัตว์อันดับวานรนั้น เมื่อ 17 ล้านปีก่อน[A] แล้วลิงวงศ์ Ponginae (ลิงอุรังอุตัง) ก็แยกออกจากสายพันธุ์เมื่อประมาณ 14 ล้านปีก่อน
จากนั้น การเดินด้วยสองเท้า (bipedalism) ซึ่งเป็นการปรับตัวพื้นฐานที่สุดของสัตว์เผ่า Hominini[B] ซึ่งเป็นสายพันธุ์ของมนุษย์ที่ลิงชิมแปนซีได้แยกออกไปแล้ว ก็เริ่มปรากฏในสัตว์สองเท้าแรกสุดในสกุล Sahelanthropus[7] (7 ล้านปีก่อน) หรือ Orrorin[8] (6.1 ล้านปีก่อน) โดยมีสกุล Ardipithecus ซึ่งเป็นสัตว์สองเท้าที่มีหลักฐานชัดเจนกว่า ตามมาทีหลัง[9] (5.8 ล้านปีก่อน) ส่วนลิงกอริลลาและลิงชิมแปนซีแยกออกจากสายพันธุ์ในช่วงเวลาใกล้ ๆ กัน คือลิงกอริลลาเมื่อ 6 ถึง 10 ล้านปีก่อน[10] และลิงชิมแปนซีเมื่อ 4 ถึง 8 ล้านปีก่อน[10] โดยอาจจะมี Sahelanthropus เป็นบรรพบุรุษสุดท้ายร่วมกันระหว่างชิมแปนซีและมนุษย์[7] สัตว์สองเท้ายุคเริ่มต้นเหล่านี้ในที่สุดก็วิวัฒนาการมาเป็นเผ่า hominini เผ่าย่อย Australopithecina (อังกฤษ: australopithecine ปกติรวมสกุล Australopithecus, Paranthropus, และในบางที่ Ardipithecus) ที่ 4.2 ล้านปีก่อน และหลังจากนั้นจึงเป็นเผ่าย่อย Hominina ซึ่งรวมเอามนุษย์สกุล โฮโม เท่านั้น[11]
มนุษย์สกุลโฮโมที่มีหลักฐานยืนยันพวกแรกที่สุดเป็นสปีชีส์ Homo habilis ซึ่งเกิดขึ้นประมาณ 2.3 ล้านปีก่อน[12][C] โดยเชื่อกันว่า สืบสายพันธุ์มาจาก homonin ในสกุล Australopithecus[13] เป็นสปีชีส์แรก ๆ ที่มีหลักฐานชัดเจนว่าใช้เครื่องมือหิน[D][15] และการปรับตัวของสายพันธุ์มนุษย์อีกอย่างหนึ่งคือ การขยายขนาดของสมอง[E] (encephalization) ก็ได้เริ่มขึ้นที่มนุษย์ยุคต้นนี้ ซึ่งมีขนาดสมองที่ประมาณ 610 ซม3 คือมีขนาดใหญ่กว่าของลิงชิมแปนซีเล็กน้อย[12] (ระหว่าง 300-500 ซม3[16]) มีนักวิทยาศาสตร์ที่เสนอว่า นี้อยู่ในช่วงเวลาที่ยีนมนุษย์ประเภท SRGAP2 มีจำนวนเป็นสองเท่าเทียบกับสัตว์เลี้ยงลูกด้วยนมอื่น ๆ[17] ซึ่งทำให้เกิดการพัฒนาของสมองกลีบหน้าได้รวดเร็วกว่าในสัตว์อื่น ๆ [F]
ต่อมา มนุษย์สปีชีส์ Homo erectus/ergaster ก็เกิดขึ้นในช่วงประมาณ 1.9 ล้านปีก่อน ที่มีปริมาตรกะโหลกศีรษะเพิ่มขึ้นเป็นสองเท่าของลิงชิมแปนซีคือ 850 ซม3[24] การขยายขนาดของสมองเช่นนี้เทียบเท่ากับมีเซลล์ประสาทเพิ่มขึ้น 125,000 เซลล์ทุกชั่วยุคคน สปีชีส์นี้เชื่อว่าเป็นพวกแรก ๆ ที่สามารถควบคุมไฟ[25] และใช้เครื่องมือหินที่มีเทคโนโลยีที่ซับซ้อนยิ่งขึ้น[15] เป็นมนุษย์สกุล Homo พวกแรกที่อพยพออกไปตั้งถิ่นฐานทั่วทวีปแอฟริกา ทวีปเอเชีย และทวีปยุโรป อาจเริ่มตั้งแต่ 1.8 ล้านปีก่อน[26] ดังนั้น การวิวัฒนาการของสายพันธุ์มนุษย์ก่อนหน้านี้ล้วนเป็นไปในแอฟริกาเท่านั้น[13]
ส่วนกลุ่มมนุษย์โบราณที่เรียกในภาษาอังกฤษว่า Archaic humans ก็เกิดวิวัฒนาการขึ้นต่อมาประมาณ 600,000 ปีก่อน[11] สืบสายพันธุ์มาจาก H. erectus/ergaster เป็นกลุ่มมนุษย์ที่อาจเป็นบรรพบุรุษของมนุษย์ปัจจุบัน โดยเฉพาะคือมนุษย์โบราณ H. heidelbergensis/rhodesiensis[27] หลังจากนั้น มนุษย์สปีชีส์ Homo sapiens ที่มีกายวิภาคปัจจุบัน (anatomically modern human) ก็เกิดขึ้นโดยมีวิวัฒนาการมาจากมนุษย์โบราณในยุคหินกลาง (แอฟริกา) คือประมาณ 300,000 ปีก่อน[G] ตามทฤษฎี "กำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้จากแอฟริกา" มนุษย์ปัจจุบันได้วิวัฒนาการในทวีปแอฟริกาแล้วจึงอพยพออกจากทวีปประมาณ 50,000-100,000 ปีก่อน (ต่างหากจากมนุษย์ในยุคก่อน ๆ) ไปตั้งถิ่นฐานแทนที่กลุ่มมนุษย์สปีชีส์ H. erectus, H. denisova, H. floresiensis และ H. neanderthalensis ในที่ต่าง ๆ ที่เป็นเชื้อสายของมนุษย์ที่อพยพออกมาจากทวีปแอฟริกาในยุคก่อน ๆ[27][28][29][30] โดยอาจได้ผสมพันธุ์กับมนุษย์โบราณก่อน ๆ เหล่านั้น[31]
หลักฐานโดยดีเอ็นเอในปี ค.ศ. 2010 บอกเป็นนัยว่า มีลำดับดีเอ็นเอหลายส่วนที่มีต้นกำเนิดจากมนุษย์โบราณ Homo neanderthalensis (อังกฤษ: Neanderthal) ในดีเอ็นเอของมนุษย์ปัจจุบันทุกเผ่าพันธุ์ที่ไม่ใช่คนแอฟริกา และว่า Neanderthal และมนุษย์โบราณสกุลอื่น ๆ เช่นที่รู้จักกันว่า Denisova hominin (อังกฤษ: Denisovan) รวม ๆ กันแล้ว อาจจะให้จีโนมเป็นส่วน 1-10% ของจีโนมมนุษย์ปัจจุบัน ซึ่งบอกเป็นนัยถึง การผสมพันธุ์กัน[H] ระหว่างมนุษย์โบราณเหล่านี้กับมนุษย์ปัจจุบัน อย่างไรก็ดี การผสมพันธุ์มีระดับค่อนข้างที่จะต่ำ และยังเป็นไปได้ว่า กรรมพันธุ์ของ Neanderthal หรือของมนุษย์โบราณอื่น ๆ ที่พบในมนุษย์ปัจจุบันอาจจะอธิบายได้โดยลักษณะสืบสายพันธุ์ (trait) ที่สืบมาจากบรรพบุรุษร่วมกันเมื่อ 500,000-800,000 มาแล้ว ไม่ใช่เพิ่งเกิดขึ้นเพราะผสมพันธุ์กันเร็ว ๆ นี้[32]
ส่วนการเปลี่ยนมามีพฤติกรรมปัจจุบัน (ดูเพิ่มที่หัวข้อ การเปลี่ยนมามีพฤติกรรมปัจจุบัน) พร้อมกับพัฒนาการของวัฒนธรรมสัญลักษณ์ (symbolic culture)[I] ภาษา และเทคโนโลยีหินแบบเฉพาะงานเริ่มขึ้นที่ประมาณ 50,000 ปีก่อนตามข้อมูลทางมานุษยวิทยา[37] แม้ว่าจะมีนักวิทยาศาสตร์บางส่วนที่เสนอว่า ความจริงเป็นการพัฒนาทางพฤติกรรมอย่างค่อย ๆ เป็นค่อย ๆ ไปในช่วงระยะเวลาที่ยาวนานยิ่งกว่านั้นที่อาจนานถึง 300,000 ปี และเริ่มมีหลักฐานแล้วว่าพฤติกรรมปัจจุบันนั้น ความจริงมีปรากฏแล้วก่อนหน้านั้น[38]
ในปัจจุบันนี้ วิวัฒนาการของมนุษย์ปัจจุบันก็ยังเป็นไปอยู่ แต่ที่ปรากฏเร็ว ๆ นี้ดูเหมือนจะจำกัดอยู่ในเรื่องภูมิต้านทานต่อโรคติดต่อโดยมาก[39] แต่เพราะไร้เหตุกดดันทางการคัดเลือกโดยธรรมชาติ หรือเพราะเหตุอื่น ๆ วิวัฒนาการของมนุษย์เร็ว ๆ นี้ โดยมากก็จะเป็นการเปลี่ยนความถี่ยีนอย่างไม่เจาะจง (genetic drift) นอกจากนั้นแล้ว ยังปรากฏอีกด้วยว่า ทั้งมนุษย์ทั้งวงศ์ลิงใหญ่แอฟริกัน (รวมกอริลลาและชิมแปนซี) ปรากฏการวิวัฒนาการที่ช้าลงจากลิงสายพันธุ์อื่น ๆ ซึ่งอาจเกิดขึ้นเพราะแต่ละชั่วอายุมีความยาวนานยิ่งขึ้น[10]
คำว่า "มนุษย์" ในบริบทของวิวัฒนาการมนุษย์ จะหมายถึงมนุษย์สกุล Homo เท่านั้น
การจัดชั้นและการใช้ชื่อในบทความ
แก้นักวิทยาศาสตร์มีความเห็นไม่เหมือนกับในการการจำแนกชั้นของสัตว์ในวงศ์ลิงใหญ่[13] แผนผังด้านล่างแสดงการจำแนกชั้นแบบหนึ่งของไพรเมต/วงศ์ลิงใหญ่ที่ยังมีชีวิตอยู่ เป็นการจำแนกชั้นที่ให้ความสำคัญกับความใกล้เคียงกันทางกรรมพันธุ์ของมนุษย์และลิงชิมแปนซี[11] โดยมีชื่อตามอนุกรมวิธาน
|
prosimian
ลิง
ลิงใหญ่
มนุษย์
ชะนี |
ส่วนการจำแนกชั้นมีดังต่อไปนี้ มีการรวมเอาสายพันธุ์มนุษย์ที่สูญพันธุ์ไว้ด้วย ชื่อแรกเป็นชื่อตามอนุกรมวิธาน ชื่อในวงเล็บเป็นชื่อทั่วไป
- วงศ์ย่อย Homininae (hominine)
- วงศ์ย่อย Gorillinae
- สกุล Gorilla (ลิงกอริลลา)
- วงศ์ย่อย Ponginae
- สกุล Pongo (ลิงอุรังอุตัง)
คำที่อาจจะมีความหมายอื่นในที่อื่น ๆ ที่ใช้บ่อย ๆ ในบทความนี้มีดังต่อไปนี้ (ตามผังด้านบน) เป็นคำที่รวมทั้งสัตว์ที่มีชีวิตอยู่และสูญพันธุ์ไปแล้ว
- วงศ์ลิงใหญ่ หรือ ลิงใหญ่ หรือ hominid หมายถึงสัตว์ในวงศ์ลิงใหญ่ หลังจากที่วงศ์ชะนีได้แยกสายพันธุ์ไปแล้ว
- homininae หรือ hominine หมายถึงสัตว์ในวงศ์ย่อย Homininae ซึ่งรวมสายพันธุ์ของมนุษย์และสายพันธุ์ของลิงชิมแปนซี หลังจากที่สายพันธุ์ของลิงกอริลลาได้แยกออกไปแล้ว
- hominini หรือ hominin (ไม่มี e ท้ายสุด) หมายถึงสัตว์ในเผ่า hominini คือสายพันธุ์มนุษย์ทั้งหมด (คือบรรพบุรุษมนุษย์ สายพันธุ์ของญาติบรรพบุรุษมนุษย์ และมนุษย์ทั้งหมด) หลังจากที่สายพันธุ์ของลิงชิมแปนซีได้แยกออกไปแล้ว
- australopithecina หรือ australopithecine ปกติหมายถึงสายพันธุ์มนุษย์สกุล Australopithecus, Paranthropus ซึ่งจัดอยู่ในเผ่าย่อย Australopithecina
- มนุษย์ หมายถึงมนุษย์สปีชีส์ต่าง ๆ ในสกุล Homo เท่านั้น ซึ่งจัดอยู่ในเผ่าย่อย Hominina
- มนุษย์โบราณ (Archaic humans) มีความหมายไม่แน่นอน ปกติรวมมนุษย์สปีชีส์ H. heidelbergensis/rhodesiensis, H. neanderthalensis และบางที่รวม H. antecessor[40]แต่โดยทั่ว ๆ ไปหมายถึงมนุษย์สปีชีส์อื่น ๆ ที่เกิดขึ้นหลังจาก 600,000 ปีก่อน รวมทั้งมนุษย์พวก Denisovans ไม่รวมมนุษย์ปัจจุบัน
- มนุษย์ปัจจุบัน หมายถึง Homo sapiens เท่านั้น โดยมักจะหมายถึง มนุษย์ที่มีกายวิภาคปัจจุบัน
หลักฐาน
แก้



หลักฐานของคำอธิบายทางวิทยาศาสตร์เกี่ยวกับวิวัฒนาการของมนุษย์มาจากงานวิทยาศาสตร์ธรรมชาติหลายสาขา (ดูรายละเอียดอื่นที่ต้นบทความ) แหล่งความรู้หลักของกระบวนการวิวัฒนาการปกติมาจากซากดึกดำบรรพ์ ซึ่งเริ่มมีการสั่งสมหลักฐานของพันธุ์มนุษย์เริ่มตั้งแต่ปี ค.ศ. 1829 (ดู "ประวัติการศึกษาสมัยดาร์วิน") แต่เริ่มตั้งแต่มีการพัฒนาด้านพันธุศาสตร์ในสาขาอณูชีววิทยาที่ต้นคริสต์ทศวรรษ 1970 (ดู "ประวัติ-การปฏิวัติทางพันธุศาสตร์") การวิเคราะห์ดีเอ็นเอก็ได้กลายมาเป็นแหล่งความรู้ที่สำคัญพอ ๆ กัน ส่วนงานศึกษาในเรื่องกำเนิดและพัฒนาการของสิ่งมีชีวิต (ontogeny) วิวัฒนาการชาติพันธุ์ (phylogeny) และโดยเฉพาะวิวัฒนาการของสิ่งมีชีวิตเชิงพัฒนาการ (evolutionary developmental biology) ของทั้งสัตว์มีกระดูกสันหลังและสัตว์ไม่มีกระดูกสันหลังได้ให้ความรู้ใหม่ ๆ พอสมควรเกี่ยวกับวิวัฒนาการของสิ่งมีชีวิตทั้งหมดรวมทั้งของมนุษย์ มีงานศึกษาเฉพาะเรื่องเกี่ยวกับกำเนิดและชีวิตของมนุษย์ ซึ่งก็คือมานุษยวิทยา (anthropology) โดยเฉพาะบรรพมานุษยวิทยา (paleoanthropology) เป็นศาสตร์ที่พุ่งความสนใจไปที่มนุษย์ก่อนประวัติศาสตร์[41]
ภายในศตวรรษที่ผ่านมาโดยเฉพาะในทศวรรษที่เพิ่งผ่าน ๆ มา ได้มีการสั่งสมหลักฐานซากดึกดำบรรพ์ (และทางอณูชีววิทยา) มากมายที่เริ่มชี้โครงสร้างการวิวัฒนาการอย่างคร่าว ๆ ของมนุษย์ปัจจุบันจากสายพันธุ์ที่แยกออกจากลิงชิมแปนซี[42] โดยที่รายละเอียดประวัติการวิวัฒนาการและการจัดชั้นของสกุลและสปีชีส์ต่าง ๆ ยังมีการเพิ่มและปรับปรุงเปลี่ยนแปลงอย่างรวดเร็วเป็นปีต่อปี[15] เพราะได้หลักฐานใหม่ ๆ ที่ช่วยยืนยันหรือปฏิเสธสมมติฐานที่มีอยู่ต่าง ๆ เพราะฉะนั้น เป็นอันหวังได้ว่า บทความจะมีข้อมูลที่ล้าหลังหลักฐานใหม่ ๆ ไปบ้าง
หลักฐานทางอณูชีววิทยา
แก้
สำหรับสัตว์ (รวมทั้งมนุษย์) ที่ยังมีชีวิตอยู่ หรือสำหรับสัตว์ที่สูญพันธุ์แล้ว (รวมทั้งสายพันธุ์ต่าง ๆ ของมนุษย์) แต่ยังหาสารอินทรีย์ที่ประกอบด้วยดีเอ็นเอได้ หลักฐานทางอณูชีววิทยานั้นสามารถให้ข้อมูลต่าง ๆ เกี่ยวกับวิวัฒนาการของมนุษย์ โดยใช้ประกอบร่วมกับข้อมูลซากดึกดำบรรพ์และข้อมูลสัตว์ที่มีอยู่ในปัจจุบัน ดังต่อไปนี้[10] คือ
- ช่วงเวลาที่สัตว์ (รวมทั้งมนุษย์) สองพันธุ์ โดยเฉพาะพันธุ์ที่ใกล้ชิดกัน
- เกิดการแยกสายพันธุ์กัน (เช่นการแยกสายพันธุ์ของมนุษย์จากลิงชิมแปนซี) หรือ
- มีบรรพบุรุษสุดท้ายร่วมกัน (เช่นมนุษย์ปัจจุบันมีบรรพบุรุษหญิงร่วมกันสุดท้ายที่ 90,000-200,000 ปีก่อน)
- ความสัมพันธ์ทางกรรมพันธุ์ระหว่างพันธุ์สัตว์ (รวมทั้งมนุษย์) ที่สามารถใช้ในการสร้างต้นไม้สายพันธุ์ (เช่นมนุษย์มีความสัมพันธ์ที่ใกล้ชิดกับลิงชิมแปนซีมากกว่าลิงกอริลลา)
- ยีนของสัตว์นั้นอาจแสดงลักษณะทางพันธุกรรมที่ปรากฏ (ซึ่งเริ่มการสั่งสมหลักฐานตั้งแต่ปี ค.ศ. 2001)
โดยที่สองข้อแรกได้ช่วยความเข้าใจในเรื่องต่าง ๆ เกี่ยวกับวิวัฒนาการมนุษย์ให้ดีขึ้นแล้ว และข้อสุดท้ายอาจมีประโยชน์ยิ่ง ๆ ขึ้นต่อ ๆ ไปในอนาคต
หลักฐานทางอณูชีววิทยา ได้ให้ข้อมูลเกี่ยวกับวิวัฒนาการมนุษย์มีตัวอย่างสำคัญ ๆ ดังต่อไปนี้
- สกุลสัตว์ที่ยังไม่สูญพันธุ์ที่ใกล้กับมนุษย์ที่สุดเป็นลิงโบโนโบ ลิงชิมแปนซี (ทั้งสองในสกุล Pan) และลิงกอริลลา (สกุล Gorilla)[11] การหาลำดับดีเอ็นเอในจีโนมของทั้งมนุษย์และลิงชิมแปนซี พบว่า มีความคล้ายคลึงกันถึงประมาณ 95-99%[44][45] เป็นความคล้ายคลึงกันที่แสดงถึงความมีสายพันธุ์เป็นพี่น้องกัน (sister taxon) หรือแม้แต่อยู่ในสกุลเดียวกัน[15]
- โดยใช้เทคนิคที่เรียกว่า molecular clock (นาฬิกาอาศัยโครงสร้างโมเลกุล) ซึ่งใช้ประเมินระยะเวลาการแยกสายพันธุ์ โดยวัดเวลาก่อนที่การกลายพันธุ์ที่ไม่เหมือนกันของสายพันธุ์สองสายพันธุ์จะสั่งสมจนมาถึงในระดับปัจจุบัน ได้มีการพบว่า การแยกสายพันธุ์ของมนุษย์และสายพันธุ์ของลิงชิมแปนซี ได้อยู่ในช่วงเวลาประมาณ 4 ถึง 8 ล้านปีก่อนซึ่งอยู่ในช่วงปลายสมัยไมโอซีน (ซึ่งเป็นส่วนปลายของยุคนีโอจีน)[10] (ดูรายละเอียดในหัวข้อ "การแยกสายพันธุ์ของวงศ์ลิงใหญ่")

"Mitochondrial Eve" อยู่เกือบบนสุด ติดกับลูกศรขาด ๆ ที่ชี้ไปที่ "Outgroup" โดยระยะห่างจากกลุ่มที่ไม่ใช่คนแอฟริกาเป็นตัวชี้ว่า สายพันธุ์ทางไมโทคอนเดรียของมนุษย์จะรวมตัวลงที่แอฟริกา
- จีโนมของมนุษย์นั้นมีทั้งส่วนที่มีการแสดงออกเป็นลักษณะสืบสายพันธุ์ (trait) และมีบางส่วนที่ไม่มีการแสดงออก ส่วนที่ไม่มีการแสดงออกสามารถใช้ในการสืบหาสายตระกูลได้ คือในส่วนที่ไม่ทำให้เกิดลักษณะสืบสายพันธุ์ การกลายพันธุ์แบบ Single-nucleotide polymorphism คือมีเบสดีเอ็นเอเปลี่ยนไปเบสเดียว จะสืบต่อไปยังลูกหลานของบุคคลนั้นทั้งหมด แต่ไม่มีในมนุษย์กลุ่มอื่น ดังนั้นสายตระกูลของบุคคลนั้นก็จะสามารถติดตามได้ ส่วนดีเอ็นเอในไมโทคอนเดรีย (mitochondrial DNA ตัวย่อ mtDNA) นั้นสืบสายมาจากมารดาเท่านั้น จึงไม่เกิดการคัดเลือกทางเพศ นอกจากนั้นแล้ว ยังเป็นดีเอ็นเอที่เกิดการกลายพันธุ์ในอัตราความถี่สูง ทำให้สามารถใช้ประเมินเวลาการแยกสายตระกูล (หรือสายพันธุ์) ได้ดี
- ผลงานวิจัยเกี่ยวกับ mtDNA หลายงานแสดงว่า หญิงที่เป็นต้นตระกูลของมนุษย์ปัจจุบันทั้งหมด ซึ่งเป็นหญิงที่เรียกว่า mitochondrial Eve (เอวาโดย mtDNA) มีชีวิตอยู่ประมาณ 90,000-200,000 ปีก่อน[46][47][48] น่าจะในแอฟริกาตะวันออก[10] ซึ่งเป็นหลักฐานที่ให้น้ำหนักกับทฤษฎีกำเนิดมนุษย์จากแอฟริกาเร็ว ๆ นี้มากขึ้น (ดูรายละเอียดเพิ่มที่หัวข้อ "การอพยพย้ายถิ่นฐานของมนุษย์")
- ผลงานวิจัยโดยจีโนมในปี ค.ศ. 2010 บอกเป็นนัยว่า มีลำดับดีเอ็นเอหลายส่วนที่มีต้นกำเนิดจากมนุษย์โบราณ Homo neanderthalensis (อังกฤษ: Neanderthal) ในดีเอ็นเอของมนุษย์ปัจจุบันทุกเผ่าพันธุ์ที่ไม่ใช่คนแอฟริกา และว่า Neanderthal และมนุษย์สกุลโฮโมสกุลอื่น ๆ เช่นกลุ่มมนุษย์โบราณที่รู้จักกันว่า Denisova hominin (อังกฤษ: Denisovan) เป็นต้นกำเนิดจีโนมถึง 1-10% ของจีโนมมนุษย์ปัจจุบัน ซึ่งบอกเป็นนัยว่า มีการผสมพันธุ์กัน[H] ระหว่างมนุษย์ปัจจุบันและมนุษย์โบราณเหล่านี้[49] (ดูรายละเอียดเพิ่มที่หัวข้อ "การผสมพันธุ์กันระหว่างมนุษย์กลุ่มต่าง ๆ")

หลักฐานจากซากดึกดำบรรพ์
แก้ซากดึกดำบรรพ์หมายถึงส่วนที่หลงเหลืออยู่ หรือร่องรอยของพืชและสัตว์ (รวมทั้งมนุษย์) ที่เกิดการเก็บรักษาไว้ในชั้นหิน ส่วนซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพ (transitional fossil) หรือเรียกอีกอย่างหนึ่งว่า สปีชีส์ในระหว่าง (intermediate species) เป็นซากดึกดำบรรพ์ของสิ่งมีชีวิตที่มีลักษณะสืบสายพันธุ์ (trait) ที่เหมือนกันกับของทั้งสัตว์กลุ่มบรรพบุรุษ และของสัตว์กลุ่มลูกหลานที่สืบเชื้อสายต่อจากกลุ่มบรรพบุรุษ[50] เป็นหลักฐานสำคัญที่สามารถใช้แสดงและติดตามวิวัฒนาการของสัตว์ เมื่อดาร์วินประกาศทฤษฎีวิวัฒนาการในปี ค.ศ. 1859 (ดูรายละเอียดในหัวข้อ "ประวัติการศึกษาสมัยดาร์วิน") หลักฐานทางซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพมีน้อยมากจนเขาได้กล่าวไว้ว่า "...เป็นข้อโต้แย้งที่เห็นได้ง่ายที่สุดที่สาหัสที่สุดต่อทฤษฎีของผม..."[51] ตั้งแต่นั้นมา ก็ได้มีการสั่งสมหลักฐานทางซากดึกดำบรรพ์มากมายที่สามารถช่วยชี้โครงสร้างกระบวนการวิวัฒนาการของมนุษย์ได้อย่างคร่าว ๆ [42]
หลักฐานทางซากดึกดำบรรพ์นั้น สามารถให้ข้อมูลโดยหลัก ๆ มีตัวอย่างดังต่อไปนี้
- ซากดึกดำบรรพ์ของสายพันธุ์มนุษย์ ซึ่งสามารถชี้กระบวนการวิวัฒนาการของมนุษย์เช่น
- ลำดับการวิวัฒนาการทางโครงสร้างจากสัตว์คล้ายลิง มาเป็นมนุษย์ปัจจุบัน (ดูหัวข้อ "ซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพ" และ "ความเปลี่ยนแปลงทางกายวิภาค")
- การวิวัฒนาการมาเป็นสัตว์สองเท้า จากสัตว์ที่อยู่บนต้นไม้ โดยอนุมานจากโครงสร้างกระดูก (ดู "ซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพ" และ "การเดินด้วยสองเท้า")
- การขยายขนาดของสมอง โดยกะโหลกศีรษะและโครงสร้างกระดูก[E] (ดูหัวข้อ "การขยายขนาดสมอง")
- ประเภทอาหารที่บริโภค โดยอนุมานจากลักษณะของใบหน้า กรามและฟัน[13][52] และโดยการวิเคราะห์อัตราส่วนของไอโซโทปในเคลือบฟัน (enamel)[53]
- เครื่องมือหิน สิ่งประดิษฐ์ วัตถุเครื่องใช้อื่น ๆ และซากดึกดำบรรพ์ของสัตว์ที่อยู่ใกล้ ๆ
- พัฒนาการของเทคโนโลยีเทียบกับกาลเวลา (ดูหัวข้อ "การใช้เครื่องมือ")
- สามารถอนุมานถึงอาหาร ความเป็นอยู่ วัฒนธรรม ความเฉลียวฉลาด และพฤติกรรมอย่างอื่น ๆ (ดูหัวข้อ "การใช้เครื่องมือ" "เครื่องมือหิน" และ "การเปลี่ยนมามีพฤติกรรมปัจจุบัน")
- ร่องรอยอื่น ๆ
- ร่องรอยของไฟ สามารถบ่งบอกการใช้และควบคุมไฟได้
- รอยเท้าสามารถบ่งการเดินด้วยสองเท้าอย่างชัดเจน (ดูหัวข้อ "รอยเท้าจากอดีต" และ "การเดินด้วยสองเท้า")
การหาอายุ
แก้โดยคร่าว ๆ แล้ว การหาอายุซากดึกดำบรรพ์แบ่งออกเป็น 3 ช่วงระยะเวลาคือ คือ
- การหาอายุจากคาร์บอนกัมมันตรังสี (radiocarbon dating) สำหรับหาอายุจากซากสิ่งมีชีวิตน้อยกว่า 40,000 ปี (หรือ 58,000 - 62,000[54])
- วิธีอื่น ๆ รวมทั้งการหาอายุจากธาตุกัมมันตรังสี (radiometric dating) แบบอื่น ๆ สำหรับหาอายุระหว่าง 40,000-100,000 ปี
- การหาอายุจากโพแทสเซียมกัมมันตรังสี (radiopotassium dating) สำหรับหาอายุมากกว่า 100,000 ปี
โดยทั่ว ๆ ไปแล้ว การหาอายุจากธาตุกัมมันตรังสีเป็นเทคนิคการหาอายุวัตถุต่าง ๆ เช่นหินหรือคาร์บอน โดยเปรียบเทียบไอโซโทปกัมมันตรังสีที่เกิดเองในธรรมชาติ กับผลิตภัณฑ์ที่เกิดจากการสลายตัวของธาตุนั้น ที่มีอัตราการสลายตัวที่ชัดเจนแล้ว[55] ส่วนการหาอายุจากคาร์บอนกัมมันตรังสีนั้นเป็นไปได้เพราะสิ่งมีชีวิตมีการบริโภคผลิตภัณฑ์ของพืชที่ได้รับคาร์บอน-14 ผ่านกระบวนการสังเคราะห์ด้วยแสงและหยุดบริโภคเมื่อหมดชีวิต ดังนั้นจึงสามารถใช้หาอายุของสารประกอบอินทรีย์จากซากสิ่งมีชีวิตโดยตรง[54] ส่วนการหาอายุจากโพแทสเซียมกัมมันตรังสีนั้นเป็นไปได้เพราะหินหลอมเหลวจากภูเขาไฟ (และเถ้า) มีธาตุนี้ ดังนั้นจึงสามารถใช้ชั้นหินเถ้าภูเขาไฟที่อยู่เหนือและใต้ซากในการประมาณอายุซากดึกดำบรรพ์ที่ต้องการได้
ตัวอย่างซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพ
แก้

hominin สปีชีส์ Australopithecus afarensis เป็นสปีชีส์ช่วงเปลี่ยนสภาพ ระหว่างบรรพบุรุษเอปที่เดินด้วยสี่เท้า กับมนุษย์ปัจจุบันที่เป็นสัตว์สองเท้า ลักษณะสายสืบพันธุ์ (trait) ของโครงกระดูกแสดงลักษณะเดินด้วยสองเท้าอย่างชัดเจน จนกระทั่งว่า นักวิชาการบางพวกเสนอว่า การเดินด้วยสองเท้าต้องเป็นวิวัฒนาการที่เกิดขึ้นก่อนสปีชีส์นี้[56] โดยทั่ว ๆ ไปแล้ว กระดูกเชิงกรานเหมือนของมนุษย์มากกว่าเอป คือ กระดูกปีกสะโพกนั้นสั้นและกว้าง ส่วนกระดูกสันหลังส่วนกระเบนเหน็บ (sacrum) ทั้งกว้างและอยู่ด้านหลังติดกับข้อสะโพก และมีหลักฐานที่ชัดเจนว่ากล้ามเนื้อเหยียดเข่า (Rectus femoris) ยึดอยู่กับกระดูกอย่างแน่น ซึ่งแสดงว่า ต้องมีอิริยาบถที่ตั้งขึ้น[56]
แม้กระดูกเชิงกรานจะไม่เหมือนกับมนุษย์ร้อยเปอร์เซนต์ (คือกว้างกว่า โดยมีกระดูกปีกสะโพกโค้งยืดออกไปข้าง ๆ) แต่ลักษณะเหล่านี้ต่างจากลิงใหญ่อื่น ๆ อย่างชัดเจน ทำให้สามารถเดินด้วยสองเท้าได้โดยระดับหนึ่ง นอกจากนั้นแล้ว กระดูกต้นขายังโค้งเข้าไปทางเข่าจากสะโพก ทำให้สามารถวางเท้าเข้าไปใกล้แนวกึ่งกลางของร่างมากขึ้น ซึ่งเป็นตัวบ่งชี้ที่ชัดเจนว่า เดินด้วยสองเท้าเป็นปกติ มนุษย์ปัจจุบัน ลิงอุรังอุตัง และลิงโลกใหม่ (ในทวีปอเมริกา) สกุล Ateles ในปัจจุบันล้วนแต่มีลักษณะแบบนี้ นอกจากนั้นแล้ว ส่วนเท้ายังมีหัวแม่โป้งหันไปทางด้านหน้า (adducted) ซึ่งทำให้การจับต้นไม้ด้วยเท้ายากหรือว่าเป็นไปไม่ได้ นอกจากการเดินแล้ว A. afarensis ยังมีฟันที่เหมือนกับมนุษย์มากกว่าเอป[57] เช่นมีขนาดเล็กกว่าเป็นต้น
ตัวอย่างรอยเท้าจากอดีต
แก้ในปี ค.ศ. 1978 ทีมของนักบรรพชีวินวิทยาแมรี ลีกคี พบรอยเท้าของ hominin เดินด้วยสองเท้าที่โบราณสถาน Laetoli ประเทศแทนซาเนีย เป็นทางยาวประมาณ 24 เมตร เป็นของ hominin 2 ตน มีอายุถึง 3.6 ล้านปีก่อน ซึ่งนักวิทยาศาสตร์เชื่อว่า เป็นรอยเท้าของ homonini สปีชีส์ Australopithecus afarensis (เหมือนกับลูซี่ข้างบน) รอยเท้าจึงทำให้หมดความสงสัยว่า hominin ที่อาจเป็นบรรพบุรุษมนุษย์พวกนี้เดินตัวตรงด้วยสองเท้าจริง ๆ [13] เพราะไม่มีรอยใช้ข้อนิ้วมือยันพื้น (เช่นในลิงชิมแปนซีและกอริลลา) นอกจากนั้นแล้ว รอยเท้ายังไม่มีรอยนิ้วแม่โป้งที่เคลื่อนไหวได้อย่างอิสระเหมือนกับของเอป แต่มีส่วนโค้งเท้าเหมือนกับมนุษย์ปัจจุบัน[58] ส่วนท่าทางการเดินดูเหมือนจะเป็นแบบสบาย ๆ
งานวิจัยในปี ค.ศ. 2007 โดยใช้การจำลองโดยคอมพิวเตอร์ที่ใช้ข้อมูลจากซากดึกดำบรรพ์ของ A. afarensis และระยะห่างของรอยเท้าที่พบแสดงว่า hominin เหล่านี้เดินด้วยความเร็วประมาณ 1.0 เมตร/วินาที หรือเร็วกว่านั้น ซึ่งเป็นความเร็วประมาณเท่ามนุษย์ปัจจุบัน[59] ผลงานวิจัยอื่น ๆ เช่นในปี ค.ศ. 2010 ยืนยันทฤษฎีว่า รอยเท้านี้เป็นของสัตว์ที่มีท่าเดินเหมือนมนุษย์[60]
วิวัฒนาการมนุษย์น่าเชื่อถือหรือไม่
แก้วิธีการศึกษาเรื่องวิวัฒนาการมนุษย์นั้นเป็นกระบวนการทางวิทยาศาสตร์ ซึ่งโดยคร่าว ๆ เป็นวงจรของการตั้งสมมติฐานและการหาหลักฐานเพื่อยืนยันหรือปฏิเสธสมมติฐานนั้น ๆ[61] เพราะฉะนั้น รายละเอียดต่าง ๆ เกี่ยวกับวิวัฒนาการมนุษย์สามารถทดสอบได้โดยหลักฐานที่มีความน่าเชื่อถือทางวิทยาศาสตร์ว่าเข้ากันกับสมมติฐานที่เป็นประเด็นศึกษาได้หรือไม่ ยกตัวอย่างเช่น ถ้าเรามีสมมติฐานว่า มนุษย์เช่นเราได้มีมาตั้งแต่กำเนิดโลก จะเป็นโลกที่เกิดขึ้นเมื่อ 6,000 ปีก่อนหรือก่อนหน้านี้ก็ตาม เมื่อขุดหาหลักฐานในที่สมควร ก็ควรจะได้ซากของมนุษย์เหมือนกับเราเริ่มต้นตั้งแต่ที่จุดกำเนิดนั้น ๆ แต่ถ้าปรากฏว่า กลับมีมนุษย์เหมือนกับเราเริ่มต้นตั้งแต่ 300,000 ปีก่อน[G] สมมติฐานว่าโลกเกิดเมื่อ 6,000 ปีก่อนพร้อมกับมนุษย์ก็ถูกปฏิเสธโดยหลักฐาน และสมมติฐานว่ามีมนุษย์เหมือนกับเราก่อนหน้า 300,000 ปีก่อน ก็ยังไม่มีหลักฐานที่จะสนับสนุน (แต่กลับมีสัตว์อื่นที่คล้ายกับเราแต่ไม่เหมือนเราก่อนหน้านั้น)
ดังนั้นประเด็นต่าง ๆ ของวิวัฒนาการมนุษย์นั้น จะเชื่อถือได้ก็โดยหลักฐานต่าง ๆ โดยเฉพาะที่ได้ผ่านการพิจารณาตามศาสตร์ต่าง ๆ ของวิทยาศาสตร์แล้วแสดงข้อสรุปเดียวกันมาแล้ว[K] ไม่ใช่เป็นเพียงแต่อาศัยความเชื่อหรือความเห็น หรือแม้แต่เพียงอาศัยสมมติฐานเท่านั้น โดยที่ยังไม่มีหลักฐานยืนยัน หรืออาจจะมีหลักฐานค้านด้วยซ้ำ[63] ประเด็นต่าง ๆ ที่มีการศึกษาอยู่บางส่วน ก็มีหลักฐานสั่งสมมามากมายจนสามารถสรุปได้อย่างชัดเจนแล้ว บางส่วนก็ยังต้องมีการสั่งสมหลักฐานเพื่อยืนยันหรือปฏิเสธสมมติฐานต่าง ๆ ต่อไป (ซึ่งหลักฐานใหม่อาจเปลี่ยนประเด็นหรือแม้แต่ข้อสรุปที่นักวิทยาศาสตร์บางส่วนมีมติเห็นด้วย - ดูตัวอย่างในหัวข้อ "การปฏิวัติทางพันธุศาสตร์" และ "ทฤษฎีอพยพสองอย่างที่ขัดแย้งกัน") และบางส่วนก็อาจจะไม่สามารถหาข้อยุติได้
ประวัติการศึกษา
แก้
ก่อนดาร์วิน
แก้คำว่า Homo ซึ่งเป็นชื่อสกุลทางชีววิทยาของมนุษย์ เป็นคำภาษาละตินที่แปลเป็นภาษาอังกฤษว่า "human" และแปลเป็นภาษาไทยว่า มนุษย์ เป็นคำที่เลือกโดยนักพฤกษศาสตร์ชาวสวีเดนคาโรลัส ลินเนียสในปี ค.ศ. 1758[64] (ผู้เริ่มเรียกสัตว์สกุลต่าง ๆ โดยใช้ทวินาม) ในระบบการจำแนกสิ่งมีชีวิตของเขา คำว่า "human" ในภาษาอังกฤษ มาจากคำภาษาละตินว่า humanus ซึ่งก็เป็นคำวิเศษณ์ของคำว่า homo[65] ลินเนียสและนักวิทยาศาสตร์อื่น ๆ ในยุคนั้นพิจารณาว่า วงศ์ลิงใหญ่เป็นสัตว์ที่ใกล้กับมนุษย์ที่สุด จากความคล้ายคลึงกันโดยสัณฐานและโดยกายวิภาค[66][67]
สมัยดาร์วิน
แก้การศึกษาทางบรรพมานุษยวิทยา (Paleoanthropology) แบบปัจจุบัน[L] ก็เริ่มขึ้นในคริสต์ศตวรรษที่ 19 เมื่อมีการค้นพบซากมนุษย์ Neanderthal ในปี ค.ศ. 1829 (ประเทศเบลเยียม) 1848 (ยิบรอลตาร์) และ 1856 (ประเทศเยอรมนี)[68] ความเป็นไปได้ว่า มนุษย์มีบรรพบุรุษเดียวกับพวกลิง ก็ปรากฏอย่างชัดเจนในปี ค.ศ. 1859 เมื่อชาลส์ ดาร์วินพิมพ์หนังสือ On the Origin of Species (กำเนิดของสปีชีส์) ซึ่งเขาได้เสนอไอเดียว่า สัตว์สปีชีส์ต่าง ๆ มีการวิวัฒนาการมาจากสปีชีส์ก่อน ๆ ที่เป็นบรรพบุรุษ หนังสือของดาร์วินไม่ได้พูดถึงวิวัฒนาการของมนุษย์โดยตรง แต่ได้กล่าวไว้เพียงแค่นี้ว่า "จะเกิดแสงสว่างในอนาคตเกี่ยวกับต้นกำเนิดและประวัติของมนุษย์"[51]
การอภิปรายถึงวิวัฒนาการมนุษย์เกิดขึ้นครั้งแรกระหว่างนักชีววิทยาโทมัส ฮักซ์ลีย์ และริชาร์ด โอเวน โดยฮักซ์ลีย์เสนอว่า มนุษย์มีวิวัฒนาการมาจากเอปโดยแสดงความคล้ายคลึงกันและความต่างกันระหว่างมนุษย์และเอป[69] โดยเฉพาะในหนังสือปี ค.ศ. 1863 ที่ชื่อว่า Evidence as to Man's Place in Nature (หลักฐานความเป็นส่วนธรรมชาติของมนุษย์) ถึงอย่างนั้น แม้นักวิทยาศาสตร์ที่สนับสนุนทฤษฎีของดาร์วิน (เช่น อัลเฟรด วอลเลซ และชาลส์ ไลเอลล์) ตอนแรกก็ไม่เห็นด้วยว่า สติปัญญาและความมีศีลธรรมของมนุษย์สามารถอธิบายได้โดยทฤษฎีการคัดเลือกโดยธรรมชาติ แม้ว่าความคิดนี้จะเปลี่ยนไปในภายหลัง[69]
ต่อมาในปี ค.ศ. 1871 ดาร์วินได้ประยุกต์ใช้ทฤษฎีวิวัฒนาการและการคัดเลือกอาศัยเพศ (sexual selection) กับมนุษย์เมื่อเขาพิมพ์หนังสือ The Descent of Man, and Selection in Relation to Sex (เชื้อสายมนุษย์ และการคัดเลือกโดยสัมพันธ์กับเพศ) [70]
ซากดึกดำบรรพ์ยุคแรก ๆ
แก้ปัญหาสำคัญเกี่ยวกับความถูกต้องของทฤษฎีวิวัฒนาการในยุคนั้นก็คือ การขาดแคลนหลักฐานในส่วนของซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพระหว่างเอปที่เป็นบรรพบุรุษกับมนุษย์ที่เป็นลูกหลาน ถึงแม้ว่านักกายวิภาคชาวดัตช์ ยูจีน ดูบัวส์ จะได้ค้นพบซากดึกดำบรรพ์ที่ปัจจุบันนี้จัดว่าเป็นส่วนของ Homo erectus ที่เกาะชวาในปี ค.ศ. 1891 (ที่นักวิทยาศาสตร์บางพวกไม่เห็นด้วยว่ากระดูกที่พบทั้งหมดมาจากบุคคลเดียวกัน[71]) นักวิทยาศาสตร์ก็ยังต้องรอจนกระทั่งถึงช่วงคริสต์ทศวรรษ 1920 กว่าจะมีการพบหลักฐานซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพเพิ่มขึ้น และเริ่มมีการสั่งสมเพิ่มขึ้นต่อ ๆ มาในทวีปแอฟริกา
ในปี ค.ศ. 1924 นักกายวิภาคชาวออสเตรเลียเรมอนด์ ดาร์ท ได้ค้นพบ hominin สปีชีส์ Australopithecus africanus ซึ่งเป็นตัวอย่างต้นแบบแรก ที่เรียกว่า Taung Child ซึ่งเป็นทารกเผ่า hominini เผ่าย่อย Australopithecina ที่ค้นพบในถ้ำแห่งหนึ่งในเมือง Taung ประเทศแอฟริกาใต้ ซากมีสภาพดีเป็นหัวกะโหลกเล็ก ๆ และเป็นรูปหล่อภายในกะโหลกศีรษะ (endocranial cast) ของสมอง ถึงแม้ว่าสมองนั้นยังเล็ก (410 ซม3) แต่ก็มีรูปร่างกลมซึ่งไม่เหมือนของลิงชิมแปนซีและกอริลลา แต่เหมือนกับของมนุษย์ปัจจุบันมากกว่า นอกจากนั้นแล้ว ตัวอย่างนั้นยังแสดงฟันเขี้ยวที่สั้น และตำแหน่งของช่องฟอราเมน แมกนัมที่เป็นหลักฐานของการเดินด้วยสองเท้า ลักษณะต่าง ๆ เหล่านี้ทำให้ดาร์ทมั่นใจว่า ทารกนั้นเป็นสัตว์บรรพบุรุษมีสองเท้าของมนุษย์ เป็นสปีชีส์ในระหว่างเอปกับมนุษย์[72]
ซากดึกดำบรรพ์จากแอฟริกาตะวันออก
แก้ในช่วงระหว่างคริสต์ทศวรรษ 1960 และ 1970 มีการค้นพบซากดึกดำบรรพ์เป็นร้อย ๆ ซาก โดยเฉพาะจากแอฟริกาตะวันออกในส่วนต่าง ๆ ของโกรกธาร Olduvai Gorge[13] และทะเลสาบ Lake Turkana[73] หัวหน้านำงานศึกษาในตอนนั้นก็คือครอบครัวลีกคี โดยตอนแรกเป็นนักบรรพชีวินวิทยาหลุยส์ ลีกคีและภรรยาคือแมรี ลีกคี และต่อจากนั้นก็เป็นบุตรชายคือริชาร์ดและลูกสะใภ้มีฟ เป็นกลุ่มที่ประสบความสำเร็จอย่างสูงสุดในการหาซากดึกดำบรรพ์ คือ จากชั้นหินซากดึกดำบรรพ์ของโกรกธาร Olduvai Gorge และทะเลสาบ Lake Turkana พวกเขาได้สั่งสมซาก hominin สาย australopithecine, มนุษย์สกุล โฮโม ยุคต้น ๆ, และแม้กระทั่งซากของ Homo ergaster/erectus[72]
การค้นพบเหล่านี้ทำให้มั่นใจได้ว่า แอฟริกาเป็นกำเนิดของมวลมนุษย์ ในช่วงคริสต์ทศวรรษ 1980 ประเทศเอธิโอเปียกลายเป็นแหล่งสำคัญใหม่ของบรรพมานุษยวิทยา[L] เมื่อ ดร. โดนัลด์ โจแฮนสัน ได้ค้นพบ "ลูซี่" ซึ่งเป็นซากดึกดำบรรพ์ของ hominin สปีชีส์ Australopithecus afarensis ที่ตอนนั้นเป็นซาก hominin สมบูรณ์ที่สุดในหมู่บ้าน Hadar ในโบราณสถาน Middle Awash ด้านเหนือของประเทศเอธิโอเปียที่เป็นเขตทะเลทราย[73] เขตนี้จะเป็นแหล่งของซากดึกดำบรรพ์ของเผ่า hominini ใหม่ ๆ มากมาย โดยเฉพาะที่ค้นพบโดยทีมของ ดร. ทิม ไวท์ ในช่วงคริสต์ทศวรรษ 1990 เช่น Ardipithecus ramidus (อาร์ดี้)[73]
การปฏิวัติทางพันธุศาสตร์
แก้การปฏิวัติทางพันธุศาสตร์เกี่ยวกับการศึกษาวิวัฒนาการของมนุษย์ก็เริ่มขึ้น เมื่อนักบรรพชีวินวิทยาชาวอเมริกันวินเซ็นต์ ซาริก และนักชีวเคมีชาวอเมริกันอัลแลน วิลสัน วัดระดับปฏิกิริยาของภูมิคุ้มกันที่เกิดจากโปรตีนแอลบูมินในน้ำเลือด (serum albumin) ในระหว่างสัตว์ที่เป็นคู่ (คือปฏิกิริยาระหว่างแอนติเจนคือแอลบูมินจากสัตว์หนึ่ง กับแอนติบอดีที่สร้างขึ้นต้านแอนติเจนนั้นในอีกสัตว์หนึ่ง) รวมทั้งมนุษย์และเอปจากทวีปแอฟริกา (คือลิงกอริลลาและลิงชิมแปนซี)[74] ระดับปฏิกิริยาสามารถกำหนดเป็นตัวเลขซึ่งเป็นตัวแทนของ "ระยะห่างทางภูมิคุ้มกัน" (immunological distance ตัวย่อ ID) ซึ่งมีสัดส่วนตามจำนวนความแตกต่างกันของกรดอะมิโนระหว่างโปรตีนกำเนิดเดียวกัน (homologous protein) ของสัตว์สปีชีส์ต่าง ๆ (ดังนั้น ระยะห่างที่ใกล้จึงหมายถึงสัตว์ที่มีความสัมพันธ์กันอย่างใกล้ชิด) โดยสร้างเส้นโค้งเทียบมาตรฐาน (calibration curve) ของ ID ระหว่างคู่สปีชีส์ เทียบกับระยะเวลาที่สายพันธุ์แยกออกจากกันตามที่มีหลักฐานแสดงโดยซากดึกดำบรรพ์ที่ชัดเจน เส้นโค้งนี้สามารถใช้เป็น molecular clock (นาฬิกาอาศัยโครงสร้างโมเลกุล) เพื่อประมาณระยะเวลาที่สายพันธุ์แยกออกจากกันระหว่างสปีชีส์คู่ที่ไม่มีซากดึกดำบรรพ์หรือมีซากที่ให้ข้อมูลได้ไม่ชัดเจน
ในผลงานวิจัยทรงอิทธิพลปี ค.ศ. 1967 ของพวกเขาในวารสาร Science ซาริกและวิลสันประมาณระยะเวลาที่สายพันธุ์แยกออกจากกันของมนุษย์และลิงใหญ่ว่าเป็น 4 ถึง 5 ล้านปีก่อน[74] ซึ่งเป็นสมัยที่คำอธิบายโดยใช้ซากดึกดำบรรพ์ประเมินระยะเวลานี้ที่ 10 ถึง 30 ล้านปีก่อน แต่ซากดึกดำบรรพ์ที่พบต่อ ๆ มา โดยเฉพาะของลูซี่ (Australopithecus afarensis) และการตีความหมายข้อมูลจากซากดึกดำบรรพ์ที่ค้นพบก่อน ๆ ใหม่ โดยเฉพาะของไพรเมตบรรพบุรุษของลิงอุรังอุตังสกุล Ramapithecus ที่มีชีวิตประมาณ 8.5-12.5 ล้านปีก่อน (สมัยไมโอซีนกลาง-ปลาย)[M] แสดงว่า ค่าประมาณที่ต่ำกว่าของซาริกและวิลสันนั้นถูกต้อง ซึ่งแสดงความถูกต้องของวิธีการวัดโดยแอลบูมิน (แต่มาถึงในสมัยปัจจุบัน ค่านั้นได้เปลี่ยนไปบ้างแล้ว ดูหลักฐานทางอณูชีววิทยา) การประยุกต์ใช้หลักของ molecular clock ได้ปฏิวัติการศึกษาเกี่ยวกับวิวัฒนาการทางโครงสร้างโมเลกุล (molecular evolution[N])
การสืบหา hominin ที่เก่าแก่ที่สุด
แก้ในช่วงคริสต์ทศวรรษ 1990 มีนักบรรพมานุษยวิทยา (paleoanthropologist) หลายทีมทั่วแอฟริกากำลังทำการค้นหาหลักฐานเกี่ยวกับเผ่า Hominini ที่เก่าที่สุดหลังจากการแยกสายพันธุ์ออกจากวงศ์ลิงใหญ่ ในปี ค.ศ. 1994 มีฟ ลีกคี ค้นพบซากของ hominin สปีชีส์ Australopithecus anamensis มีอายุประมาณ 4 ล้านปี แต่ว่าการค้นพบนี้ถูกทำให้อับแสงด้วยการค้นพบ hominin สปีชีส์ Ardipithecus ramidus ของทิม ไวท์ ในปีเดียวกัน ซึ่งมีประมาณอายุที่ 4.4 ล้านปี[72]
ในปี ค.ศ. 2000 นักบรรพมานุษยวิทยาชาวฝรั่งเศสมาร์ติน พิกฟอร์ด และบริกีต์ เซนูต์ ค้นพบ hominin ที่เดินด้วยสองเท้า มีอายุ 6 ล้านปีในบรรพชีวินสถาน Tugen Hills ของประเทศเคนยา ซึ่งพวกเขาได้ตั้งชื่อสปีชีส์ว่า Orrorin tugenensis[13] ในปี ค.ศ. 2001 ทีมนักวิจัยนำโดยนักบรรพชีวินวิทยาชาวฝรั่งเศสไมเคิล บรูเนต์ ค้นพบกะโหลกศีรษะของสัตว์วงศ์ลิงใหญ่ (หรืออาจจะเป็นเผ่า hominini) สปีชีส์ Sahelanthropus tchadensis อายุ 7 ล้านปี[75] ซึ่งบรูเนต์อ้างว่า เป็นสัตว์สองเท้า และดังนั้นจึงจัดอยู่ในเผ่า hominini[7]
การย้ายถิ่นฐานของมนุษย์
แก้
นักมานุษยวิทยาในช่วงคริสต์ทศวรรษ 1980 ไม่มีมติร่วมกันเกี่ยวกับรายละเอียดการอพยพย้ายถิ่นฐานของมนุษย์สกุลโฮโม ต่อจากนั้น ก็เริ่มมีการใช้วิธีการทางกรรมพันธุ์เพื่อตรวจสอบและแก้ประเด็นปัญหาต่าง ๆ เหล่านี้
การอพยพของมนุษย์ยุคต้นก็เริ่มต้นขึ้นเมื่อมนุษย์ H. ergaster/erectus เป็น Homo รุ่นแรกที่อพยพออกจากแอฟริกาผ่านช่องทางที่เรียกว่า "Levantine corridor" (ช่องทางเลแวนต์) โดยผ่านเขตจะงอยแอฟริกา ไปทางยูเรเชีย เริ่มตั้งแต่ 1.8 ล้านปีก่อน[26] หลังจากนั้นก็ตามด้วย H. antecessor ไปทางยุโรปเมื่อ 800,000 ปีก่อน แล้วตามด้วย H. heidelbergensis ซึ่งเป็นสปีชีส์ที่น่าจะเป็นบรรพบุรุษของทั้งมนุษย์ปัจจุบัน (โดยสายที่อยู่ในแอฟริกา) และของ Neanderthal (โดยสายที่อยู่ในยูเรเชีย) เมื่อ 600,000 ปีก่อน[76]
ทฤษฎีอพยพสองอย่างที่ขัดแย้งกัน
แก้"ทฤษฎีกำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้จากแอฟริกา" (Recent African origin of modern humans หรือสั้น ๆ ว่า Recent African origin) เสนอว่า มนุษย์ปัจจุบันคือ H. sapiens เกิดเป็นสปีชีส์ใหม่ในทวีปแอฟริกาเมื่อเพียงเร็ว ๆ นี้ (ประมาณ 300,000 ปีก่อน[G]) จากบรรพบุรุษในแอฟริกา ไม่ใช่จากพวกมนุษย์ที่อพยพออกมาแล้ว และมีการอพยพของมนุษย์สปีชีส์นี้ผ่านทวีปยูเรเชียไปตั้งถิ่นฐานในที่ต่าง ๆ เป็นการทดแทนมนุษย์สกุลโฮโม อื่น ๆ เกือบสิ้นเชิง คริส สตริงเกอร์ และปีเตอร์ แอนดรูส์ ได้พัฒนาแบบจำลองทางวิทยาศาสตร์นี้ในปี ค.ศ. 1988[77] ซึ่งพร้อมกับแบบจำลองของการอพยพออกจากแอฟริกาตะวันออก เป็นแบบกำเนิดมนุษย์ที่หมู่วิทยาศาสตร์ยอมรับมากที่สุดจนถึงปี ค.ศ. 2006[78][79][28]
โดยเปรียบเทียบกัน "สมมติฐานวิวัฒนาการภายในหลายเขต" (Multiregional Evolution) เสนอว่ามนุษย์สกุลโฮโม มีเพียงสปีชีส์เดียวที่มีกลุ่มประชากรต่าง ๆ ที่มีการติดต่อกันตั้งแต่ต้น ไม่ใช่มีสปีชีส์ต่าง ๆ กัน และวิวัฒนาการของมนุษย์ก็เกิดขึ้นทั่วโลกอย่างสืบเนื่องกันตลอด 2-3 ล้านปีที่ผ่านมา นักบรรพมานุษยวิทยาชาวอเมริกันมิลฟอร์ด วอลปอฟฟ์ ได้เสนอแบบจำลองนี้ในปี ค.ศ. 1988[80]
"Mitochondrial Eve" อยู่เกือบบนสุด ติดกับลูกศรขาด ๆ ที่ชี้ไปที่ "Outgroup" โดยระยะห่างจากกลุ่มที่ไม่ใช่คนแอฟริกาเป็นตัวชี้ว่า สายพันธุ์ทางไมโทคอนเดรียของมนุษย์จะรวมตัวลงที่แอฟริกา
ทฤษฎีกำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้มีน้ำหนักกว่า
แก้ภายในสองทศวรรษที่ผ่านมา มีการพบซากดึกดำบรรพ์ Homo sapiens ที่เก่าแก่ที่สุดในโลกในแอฟริกา คือ
- ในปี ค.ศ. 2017 มีการค้นพบกระดูกกะโหลกศีรษะของบรรพบุรุษมนุษย์ปัจจุบันจากโบราณสถาน Jebel Irhoud ประเทศโมร็อกโก อายุประมาณ 315,000 ปี[G]
- ในปี ค.ศ. 1997 (เผยแพร่ 2003) มีการพบกะโหลกศีรษะ 3 กะโหลกในเขต Middle Awash เอธิโอเปีย อายุประมาณ 160,000 ปี[81]
- ซากดึกดำบรรพ์ที่เรียกว่า Omo remains ขุดได้จากเขตใกล้กับ Middle Awash มีอายุประมาณ 195,000 ปี[82]
เหล่านี้เป็นหลักฐานส่วนหนึ่งที่อาจแสดงว่า มนุษย์ปัจจุบันมีกำเนิดมาจากทวีปแอฟริกา[31] นอกจากนั้นแล้ว งานวิจัยโดยใช้ haplogroup ใน Y-DNA และ mtDNA ต่อมา ยังสนับสนุนทฤษฎีกำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้โดยมาก[83]
ความก้าวหน้าในการหาลำดับดีเอ็นเอ โดยเฉพาะในดีเอ็นเอของไมโทคอนเดรีย (mtDNA) และต่อจากนั้น ในดีเอ็นเอของโครโมโซม Y (Y-DNA) ได้ช่วยสร้างความเข้าใจเกี่ยวกับกำเนิดมนุษย์ให้ดีขึ้น คือ การหาลำดับของทั้ง mtDNA และ Y-DNA ที่ได้ตัวอย่างมาจากคนพื้นเมืองในที่ต่าง ๆ ได้แสดงข้อมูลเกี่ยวกับบรรพบุรุษที่สืบมาทั้งจากทางมารดาและทางบิดา คือได้พบว่า มนุษย์ปัจจุบันทั้งหมดสืบสายโดยไม่ขาดตอนมาจากหญิงคนเดียวกัน[10] ระหว่าง 90,000-200,000 ปีก่อน[46][47][48] (ผู้เรียกว่า mitochondrial Eve) และมาจากชายคนเดียวกันผู้น่าจะอยู่ในแอฟริกากลาง-ตะวันตกเฉียงเหนือ[84] ระหว่าง 180,000-338,000 ปีก่อน[85][86] (ผู้เรียกว่า Y-DNA Adam)
นอกจากนั้นแล้ว ยังมีงานวิเคราะห์ที่ได้แสดงระดับความหลากหลายของรูปแบบดีเอ็นเอทั่วทวีปแอฟริกาที่สูงกว่านอกทวีป ซึ่งเข้ากับไอเดียว่า แอฟริกาเป็นแหล่งกำเนิดของมารดาที่เป็นต้นแบบของ mtDNA (mitochondrial Eve) และบิดาที่เป็นต้นแบบของ Y-DNA (Y-chromosomal Adam)[P] ข้อมูลทางกรรมพันธุ์เช่นนี้มีการตีความหมายว่า สนับสนุนทฤษฎีกำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้จากแอฟริกา
การผสมพันธุ์กันระหว่างมนุษย์กลุ่มต่าง ๆ
แก้อย่างไรก็ดี ก็ยังมีหลักฐานเกี่ยวกับการผสมพันธุ์[H] กันระหว่างมนุษย์ปัจจุบันกับสปีชีส์ต่าง ๆ ของกลุ่มมนุษย์โบราณอีกด้วย[32][49] คือ ในปี ค.ศ. 2010 การหาลำดับดีเอ็นเอของ Neanderthal (Homo neanderthalensis) และของ Denisovan (Denisova hominin) พบว่า มีการผสมพันธุ์ของมนุษย์ปัจจุบันกับมนุษย์สปีชีส์อื่น ๆ คือ มนุษย์ปัจจุบันนอกทวีปแอฟริกามีอัลลีลของ Neanderthal ประมาณ 1-3% ในจีโนม ของตน และชาวเมลานีเซียยังมีอัลลีลของ Denisovan อีก 3.5% ) เทียบกับมนุษย์ปัจจุบันอื่น คือ HLA haplotype ที่มีกำเนิดจาก Neanderthal และ Denisovan มีทั้งในคนยูเรเชีย และในคนโอเชียเนียในปัจจุบัน แต่ในคนแอฟริกาไม่มี นอกจากนั้นแล้ว งานวิจัยอื่นในปี ค.ศ. 2011 และ 2012 ยังพบด้วยอีกว่า กลุ่มชนแอฟริกาใต้สะฮารามีเชื้อสายมาจากกลุ่มมนุษย์โบราณที่ยังไม่รู้จัก โดยปัจจุบันสูญพันธุ์ไปหมดแล้ว[32]
แต่ว่า ผลงานทางพันธุกรรมเหล่านี้ไม่ได้ขัดแย้งกับทฤษฎีกำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้ ยกเว้นแต่นัยที่ตีความหมายอย่างแคบที่สุด (คือไม่มีการผสมพันธุ์ระหว่างมนุษย์ปัจจุบันและมนุษย์โบราณ) คือมีนักวิทยาศาสตร์บางพวกที่ชอบใจทฤษฏีที่รวมเอาองค์ประกอบของทฤษฎีที่ขัดแย้งกันทั้งสองเพราะว่าสามารถอธิบายข้อมูลได้ดีที่สุด คือว่า ไม่ได้เกิดสปีชีส์ของมนุษย์ใหม่จริง ๆ ในแอฟริกา (ซึ่งทำให้การผสมพันธุ์กับมนุษย์รุ่นก่อน ๆ เป็นไปได้) แต่ว่า มีมนุษย์รุ่นใหม่ที่มีกายวิภาคปัจจุบันที่อพยพออกไปจากแอฟริกาแล้วเข้าไปตั้งถิ่นฐานแทนที่มนุษย์รุ่นก่อน ๆ โดยในบางที่อาจจะมีการผสมพันธุ์กันกับมนุษย์รุ่นก่อน ๆ แม้ว่าจะมีน้อย[31][13] คือ ผลงานทางกรรมพันธุ์ (2010, 2011, 2012) สามารถอธิบายได้ว่า หลังจากการฟื้นตัวจากคอคอดประชากร (population bottleneck) ที่อาจมีสาเหตุมาจากมหันตภัยภูเขาไฟทะเลสาบโตบาประมาณ 75,000 ปีก่อน มนุษย์กลุ่มค่อนข้างเล็กได้ผสมสายพันธุ์กับ Neanderthal ซึ่งน่าจะเกิดขึ้นที่ตะวันออกกลาง หรือแอฟริกาเหนือก่อนที่จะอพยพออกจากทวีปแอฟริกา และลูกหลานของมนุษย์กลุ่มนี้ซึ่งก็ยังเป็นเชื้อสายคนแอฟริกาโดยมาก ก็ได้อพยพไปตั้งถิ่นฐานในที่ต่าง ๆ ทั่วโลก และมีส่วนหนึ่งที่ผสมสายพันธุ์กับ Denisovan น่าจะที่เอเชียอาคเนย์ ก่อนที่จะไปถึงเมลานีเซีย[32]
อีกอย่างหนึ่ง การผสมพันธุ์กับมนุษย์มนุษย์โบราณมีระดับค่อนข้างที่จะต่ำ (1-10%) และยังมีงานวิจัยอื่นหลังจาก ปี ค.ศ. 2010 ที่เสนอว่า กรรมพันธุ์ของมนุษย์โบราณอื่น ๆ ที่พบในมนุษย์ปัจจุบันอาจจะอธิบายได้โดยลักษณะสืบสายพันธุ์ (trait) ที่สืบมาจากบรรพบุรุษร่วมกันเมื่อ 500,000-800,000 มาแล้ว ไม่ใช่เพิ่งเกิดขึ้นเพราะผสมพันธุ์กันเร็ว ๆ นี้[32]
แบบจำลองมีการอพยพหลายครั้ง
แก้ถึงกระนั้น ทฤษฎีกำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้ ก็ยังมีทฤษฎีย่อยต่าง ๆ กันอีกว่า เป็นการอพยพเพียงครั้งเดียว หรือหลายครั้ง แบบจำลองอพยพหลายครั้ง (multiple dispersal model) รวมเอาทฤษฎีอพยพไปทางทิศใต้ (Southern Dispersal theory) เข้าไว้ด้วย[87] ซึ่งเริ่มได้รับหลักฐานสนับสนุนทางกรรมพันธุ์ ทางภาษา และทางโบราณคดี เป็นทฤษฎีที่บอกว่า มีการอพยพของมนุษย์ปัจจุบันจากเขตติดทะเลคือจากจะงอยแอฟริกา (คือจากแอฟริกาตะวันออกเฉียงเหนือ หรือแหลมโซมาลี) ประมาณ 70,000 ปีก่อนเป็นกลุ่มแรก กลุ่มนี้ไปตั้งถิ่นฐานอยู่ในเอเชียอาคเนย์และโอเชียเนีย ซึ่งอธิบายโบราณสถานในเขตเหล่านี้ ที่เก่าแก่ยิ่งกว่าในเขตเลแวนต์ (เขตฝั่งทะเลเมดิเตอร์เรเนียนทิศตะวันออก) แต่หลักฐานของการอพยพตั้งถิ่นฐานของมนุษย์กลุ่มแรกโดยมากน่าจะถูกทำลายโดยระดับน้ำทะเลที่สูงขึ้นหลังจากยุคน้ำแข็งแต่ละยุคสิ้นสุดลง เพราะเป็นการตั้งถิ่นฐานในเขตฝั่งทะเลที่ถูกน้ำท่วมหลังจากน้ำแข็งละลายแล้วทะเลสูงขึ้น[87]
ส่วนมนุษย์กลุ่มที่สองออกจากทวีปแอฟริกาข้ามคาบสมุทรไซนายเข้าไปในเอเชีย แล้วทำการตั้งถิ่นฐานโดยมากในทวีปยูเรเชีย กลุ่มนี้อาจจะมีเทคโนโลยีเครื่องมือที่ทันสมัยกว่า[87] จึงต้องอาศัยแหล่งอาหารทางทะเลน้อยกว่ากลุ่มแรก แต่ว่าแบบจำลองอพยพหลายครั้งมีหลักฐานคัดค้านจากงานวิจัยในปี ค.ศ. 2005 ที่แสดงว่า กลุ่มประชากรในทวีปยูเรเชีย (กลุ่มสอง) และในเอเชียอาคเนย์และโอเชียเนีย (กลุ่มแรก) ล้วนแต่มี mtDNA เชื้อสายเดียวกัน ซึ่งเป็นข้อมูลที่สนับสนุนว่ามีการอพยพเพียงสายเดียวออกจากแอฟริกาตะวันออกที่เป็นเหตุกำเนิดของประชากรนอกแอฟริกาทั้งหมด[88]

ความเปลี่ยนแปลงทางกายวิภาค
แก้วิวัฒนาการของมนุษย์สามารถกำหนดได้โดยความเปลี่ยนแปลงทางสัณฐาน (morphological) ทางพัฒนาการ (developmental) ทางสรีรภาพ (physiological) และทางพฤติกรรม ที่เกิดขึ้นตั้งแต่การแยกออกจากสายพันธุ์ของลิงชิมแปนซี การปรับตัวที่สำคัญที่สุดก็คือการเดินด้วยสองเท้า (bipedalism), การขยายขนาดของสมอง (encephalization[E]), ช่วงการพัฒนาและการเติบโตที่ยาวนานขึ้น (ทั้งในท้องและในวัยเด็ก) และความแตกต่างระหว่างเพศ (sexual dimorphism) ที่ลดลง แต่ความสัมพันธ์ต่อกันและกันของความเปลี่ยนแปลงต่าง ๆ เหล่านี้ยังเป็นเรื่องที่ยังไม่มีข้อยุติ[89] การเปลี่ยนแปลงทางสัณฐานอื่น ๆ ที่สำคัญรวมทั้งวิวัฒนาการของการจับวัตถุได้อย่างแม่นยำและมีกำลัง ซึ่งเกิดขึ้นครั้งแรกใน Homo ergaster/erectus[90]
การเดินด้วยสองเท้า
แก้
การเดินด้วยสองเท้า (bipedalism) เป็นการปรับตัวขั้นพื้นฐานของสัตว์เผ่า hominini[B] และพิจารณาว่าเป็นเหตุของการเปลี่ยนแปลงทางโครงสร้างกระดูกต่าง ๆ ที่ hominin ทุก ๆ สกุลมี[15]
เผ่า hominini ที่เดินด้วยสองเท้าที่เก่าแก่ที่สุดพิจารณาว่าเป็นสกุล Sahelanthropus[7] หรือไม่ก็สกุล Orrorin[8] โดยมีสกุล Ardipithecus ซึ่งเป็นสัตว์สองเท้าที่มีหลักฐานชัดเจนกว่า ตามมาทีหลัง[9] ส่วนลิงที่เดินโดยใช้ข้อนิ้วมือคือลิงกอริลลาและลิงชิมแปนซีแยกออกจากสายพันธุ์ในเวลาใกล้ ๆ กัน โดยอาจจะมีสกุล Sahelanthropus[7] เป็นบรรพบุรุษสุดท้ายร่วมกันกับมนุษย์
สัตว์สองเท้ายุคเริ่มต้นเหล่านี้ในที่สุดก็วิวัฒนาการเป็นเผ่า hominini เผ่าย่อย australopithecina (อังกฤษ: australopithecine ปกติรวมเอาสกุล Australopithecus และ Paranthropus) และหลังจากนั้นจึงเป็นเผ่าย่อย hominina ซึ่งรวมเอามนุษย์สกุล โฮโม เท่านั้น มีทฤษฎีหลายทฤษฎีถึงประโยชน์ในการปรับตัวใช้สองเท้า[13][15] เป็นไปได้ว่า การเดินด้วยสองเท้าได้รับการคัดเลือกโดยธรรมชาติ เพราะว่า ทำให้มือเป็นอิสระในการจับสิ่งของและในการถืออาหาร, เป็นการประหยัดพลังงานในขณะเดินทาง, ทำให้สามารถวิ่งได้และล่าสัตว์ได้ไกล ๆ, ทำให้เห็นได้ดีขึ้น และช่วยป้องกันการเกิดความร้อนเกินโดยลดเนื้อที่ผิวที่ถูกแสงอาทิตย์ การปรับตัวทั้งหมดนี้ก็อาจเพื่อมีสภาพที่เหมาะสมยิ่งขึ้นกับสิ่งแวดล้อมที่เปลี่ยนแปลงไปที่นักวิทยาศาสตร์มีสมมติฐานต่าง ๆ เป็นต้นว่า[13]
- มีป่าลดลง มีทุ่งหญ้าเพิ่มขึ้น โดยเฉพาะในแอฟริกาตะวันออก ทำให้จำเป็นในการเดินด้วยสองเท้า (savanna hypothesis)
- มีป่าที่ผสมกับทุ่งหญ้า เปิดโอกาสให้หากินได้ทั้งบนต้นไม้และบนพื้น (mosaic hypothesis)
- มีสิ่งแวดล้อมและภูมิอากาศที่เปลี่ยนไปอย่างรวดเร็ว ทำให้จำเป็นที่จะต้องปรับตัวให้เข้ากับสิ่งแวดล้อมหลาย ๆ อย่างได้ (variability hypothesis)
โดยกายวิภาค การเดินด้วยสองเท้าต้องประกอบพร้อมกับความเปลี่ยนแปลงในโครงสร้างกระดูก ซึ่งไม่ใช่เพียงแค่ขาที่ยาวขึ้นและเชิงกรานที่เปลี่ยนรูปร่างไปเท่านั้น แต่กับส่วนอื่น ๆ เช่นกระดูกสันหลัง เท้ากับข้อเท้า และกะโหลกศีรษะด้วย[15] คือ กระดูกต้นขาเกิดวิวัฒนาการโดยโค้งเข้ามาทางศูนย์กลางความโน้มถ่วง เข้ามาแนวกลางด้านตั้งของร่างกาย หัวข้อเข่าและข้อเท้าก็แข็งแกร่งยิ่งขึ้นเพื่อรองรับน้ำหนักที่เพิ่มขึ้น ส่วนกระดูกสันหลังก็เปลี่ยนไปเป็นรูป S เพื่อที่แต่ละข้อจะรองรับน้ำหนักมากขึ้นเมื่อยืน และกระดูกสันหลังระดับเอว (lumbar vertebrae) ก็สั้นลงและกว้างขึ้น ส่วนที่เท้า หัวแม่โป้งก็หันไปทางเดียวกันกับนิ้วเท้าอื่น ๆ ที่สั้นลงเพื่อช่วยในการเดินไปข้างหน้า กระดูกเท้าก็เพิ่มส่วนโค้งในทางยาว แขนทั้งส่วนต้นส่วนปลายก็สั้นลงเทียบกับขาเพื่อทำให้วิ่งสะดวกยิ่งขึ้น ช่องฟอราเมน แมกนัมซึ่งเป็นทางออกของไขสันหลังที่กะโหลกศีรษะ ก็ย้ายไปอยู่ทางด้านล่างของกะโหลกเยื้องไปทางด้านหน้า[91] เทียบกับของลิงใหญ่ที่เยื้องไปทางด้านหลัง[11]

แต่ความเปลี่ยนแปลงที่สำคัญที่สุดอยู่ที่กระดูกเชิงกราน โดยที่กระดูกปีกสะโพกที่ก่อนหน้านี้ยื่นยาวไปด้านล่าง เกิดสั้นลงและกว้างขึ้น (ดูรูป) ซึ่งจำเป็นเพื่อรองรับอวัยวะภายในขณะยืนและเดิน ดังนั้น hominin ที่เดินด้วยสองเท้าจึงมีกระดูกเชิงกรานที่สั้นกว่า แต่กว้าง มีรูปร่างคล้ายชาม[13] จุดอ่อนก็คือ ช่องคลอดในหญิงสายพันธุ์มนุษย์มีขนาดเล็กกว่าเอปที่เดินใช้หลังกระดูกข้อนิ้วมือ แม้ว่าจะมีการขยายใหญ่ขึ้นอีกในบางสกุลโดยเฉพาะของมนุษย์ปัจจุบัน (เมื่อเทียบกับของ australopithecine) เพื่อให้ทารกที่มีกะโหลกศีรษะใหญ่ขึ้นผ่านออกมาได้ แต่ขนาดที่ใหญ่ขึ้นจำกัดอยู่ที่ด้านบนของเชิงกรานเท่านั้น เพราะว่า ความเปลี่ยนแปลงมากกว่านี้สามารถเป็นอุปสรรคแก่การเดินด้วยสองเท้าได้[96]
เชิงกรานที่สั้นลงและช่องคลอดที่เล็กลงเป็นการวิวัฒนาการเพื่อให้เดินด้วยสองเท้าได้ แต่มีผลสำคัญต่อการคลอดลูกในมนุษย์ปัจจุบัน ซึ่งยากกว่าในไพรเมตอื่น ๆ คือ ในขณะออกจากท้องแม่ เพราะว่าส่วนต่าง ๆ ของช่องผ่านเชิงกรานมีขนาดต่าง ๆ กัน ศีรษะของทารกจะต้องเยื้องไปทางตะโพกของแม่ด้านใดด้านหนึ่งเมื่อเริ่มเข้าไปในช่องคลอด และจะต้องหมุนไปในช่วงต่าง ๆ กันประมาณ 90 องศาก่อนที่จะออก (ดูรูป)[97] ช่องคลอดที่เล็กลงกลายเป็นอุปสรรคเมื่อขนาดสมองเริ่มขยายใหญ่ขึ้นในมนุษย์ยุคต้น ๆ มีผลทำให้มีระยะการตั้งครรภ์ที่สั้นลง ซึ่งอาจเป็นเหตุผลที่มนุษย์ให้กำเนิดทารกที่ยังไม่สมบูรณ์ ผู้ไม่สามารถเดินได้ก่อนวัย 12 เดือนและมีการเจริญเติบโตที่ยืดเยื้อ เมื่อเทียบกับไพรเมตอื่น ๆ ซึ่งสามารถเดินได้ในวัยเด็กกว่า[98]
สมองที่ต้องมีการพัฒนาในระดับที่สูงขึ้นหลังคลอด และการต้องอาศัยแม่มากขึ้นของเด็ก มีผลอย่างสำคัญต่อวงจรสืบพันธ์ของหญิง[ต้องการอ้างอิง] และสำหรับนักวิชาการบางท่าน ต่อการมีคู่ครองคนเดียวที่ปรากฏบ่อยครั้งในมนุษย์เมื่อเทียบกับสกุลวงศ์ลิงใหญ่อื่น ๆ [99][100] แม้ว่า จะมีนักวิชาการท่านอื่นที่มีความเห็นว่า การมีคู่ครองคนเดียวไม่เคยเป็นส่วนของการสืบพันธุ์หลักใน hominin[101] และนอกจากมีระยะเวลาที่ยืดเยื้อก่อนจะถึงวัยเริ่มเจริญพันธุ์แล้ว ยังมีการเกิดวัยหมดระดูอีกด้วย โดยมีสมมติฐานหนึ่งที่ชี้ว่า หญิงที่สูงวัยขึ้นสามารถสืบสายพันธ์ของตนได้ดีกว่าถ้าช่วยดูแลลูกของลูกสาว ถ้าเทียบกับต้องดูแลลูกของตนที่มีเพิ่มอีก[102]
การขยายขนาดสมอง
แก้มนุษย์สปีชีส์ต่าง ๆ มีสมองที่ใหญ่กว่าไพรเมตประเภทอื่น ๆ ซึ่งในมนุษย์ปัจจุบันมีขนาดเฉลี่ย 1,330 ซม3 ใหญ่กว่าของลิงชิมแปนซีหรือลิงกอริลลามากกว่าสองเท่า[103] การขยายขนาดสมอง[E] เริ่มขึ้นที่มนุษย์สกุล Homo habilis ซึ่งมีขนาดสมองที่ประมาณ 610 ซม3 เป็นขนาดที่ใหญ่กว่าลิงชิมแปนซีเล็กน้อย (ระหว่าง 300-500 ซม3[16]) ตามมาด้วย Homo ergaster/erectus ที่ขนาดเฉลี่ย 850 ซม3[24] ไปสุดที่ Neanderthal ที่ขนาดเฉลี่ย 1,500 ซม3[104] ซึ่งใหญ่กว่าของมนุษย์ปัจจุบันเสียอีก
นอกจากนั้นแล้ว รูปแบบการพัฒนาของสมองหลังคลอด ก็ยังแตกต่างจากเอปประเภทอื่น ๆ อีกด้วย ทำให้มนุษย์สามารถศึกษาเรียนรู้ทางด้านสังคมและภาษาเป็นระยะเวลานานในวัยเด็ก อย่างไรก็ดี ความแตกต่างโดยโครงสร้างสมองของมนุษย์เทียบกับเอป อาจจะมีความสำคัญยิ่งกว่าความแตกต่างโดยขนาด[103]
ขนาดที่ขยายใหญ่ในแต่ละเขตของสมองไม่เท่ากัน คือ สมองกลีบขมับ ซึ่งมีศูนย์ประมวลผลทางภาษา ได้ขยายใหญ่ขึ้นมากกว่าเขตอื่น ๆ และก็เป็นจริงด้วยสำหรับ prefrontal cortex (คอร์เทกซ์กลีบหน้าผากส่วนหน้า) ซึ่งมีบทบาทเกี่ยวกับการตัดสินใจที่ซับซ้อนและการควบคุมพฤติกรรมในสังคม (ดู executive functions)[103] การขยายขนาดของสมองเชื่อว่ามีความสัมพันธ์กับการมีเนื้อสัตว์เพิ่มเป็นอาหาร[103] หรือกับการหุงอาหาร[105] และมีการเสนอว่า มนุษย์มีเชาวน์ปัญญาที่สูงขึ้นเป็นการตอบสนองต่อความจำเป็นในการแก้ปัญหาทางสังคม (เป็นทฤษฎีที่เรียกว่า Social brain hypothesis) เมื่อสังคมมนุษย์มีความซับซ้อนมากยิ่งขึ้น[103]
ความแตกต่างระหว่างเพศ
แก้ระดับความแตกต่างระหว่างเพศ (sexual dimorphism) ที่ลดลงในมนุษย์ จะเห็นได้ในเพศชายโดยหลัก คือขนาดที่เล็กลงของฟันเขึ้ยวเทียบกับเอปประเภทอื่น ๆ ขนาดที่เล็กลงของสันคิ้ว[13] และความแข็งแรงโดยทั่วไปที่ลดลง
ความแตกต่างที่สำคัญทางกายภาพอีกอย่างหนึ่งทางเพศของมนุษย์ก็คือ การมีช่วงตกไข่ที่ซ่อนเร้นในเพศหญิง[102] คือมนุษย์เป็นเอปประเภทเดียวที่เพศหญิงตั้งครรภ์ได้ตลอดทั้งปี โดยไม่มีสัญญาณพิเศษที่แสดงออกทางร่างกาย (เช่นความบวมขึ้นของอวัยวะเพศเมื่ออยู่ในช่วงตกไข่) แม้ว่าจะมีงานวิจัยเร็ว ๆ นี้ที่แสดงว่า หญิงมักจะมักมีความคิดและอารมณ์ทางเพศเพิ่มขึ้นก่อนที่จะตกไข่[106]
อย่างไรก็ดี มนุษย์ก็ยังมีความแตกต่างระหว่างเพศในระดับหนึ่ง เช่นรูปแบบการแพร่กระจายของขนและไขมันใต้ผิว และขนาดทั่วไปของร่างกาย โดยที่ชายมีขนาดใหญ่กว่าหญิงประมาณ 15% (ข้อมูลของสหรัฐอเมริกา)[107] ความเปลี่ยนแปลงเหล่านี้โดยองค์รวมถือกันว่าเป็นผลจากการเพิ่มความสำคัญของการมีชีวิตคู่ (pair bonding) เป็นการแก้ปัญหาที่พ่อแม่ต้องเลี้ยงลูกเป็นระยะเวลายาวนานยิ่งขึ้น[ต้องการอ้างอิง]
การปรับตัวทางกายภาพอื่น ๆ
แก้
มีความเปลี่ยนแปลงอื่น ๆ ที่เป็นลักษณะเฉพาะของวิวัฒนาการในมนุษย์ รวมทั้ง
- โครงสร้างของมือ ข้อมือ และนิ้วที่ทำให้จับสิ่งของได้อย่างมีกำลังยิ่งขึ้น อย่างละเอียดละออยิ่งขึ้น[108] อย่างมีอิสระมากขึ้น ซึ่งมีการอ้างว่า เป็นการปรับตัวเพื่อทำและใช้เครื่องมือ[11]
- โครงสร้างของแขนที่ไม่ได้ใช้รับน้ำหนักจึงมีกระดูกที่ตรงกว่า สั้นกว่า มีกล้ามเนื้อที่มีพลังน้อยกว่า แต่สามารถเคลื่อนที่ไปอย่างมีอิสระกว่าและได้ในระดับความเร็วต่าง ๆ กันมากกว่า[11]
- มีทางเดินอาหารที่สั้นกว่าและเล็กกว่า[11]
- ร่างกายมีขนน้อยลง และผมมีการงอกที่ช้าลง[10]
- การเปลี่ยนรูปของแนวฟันจากรูปค่อนข้างเหลี่ยม (เหมือนอักษรโรมัน U) เป็นรูปโค้ง (เหมือนพาราโบลา)[13]
- การยื่นออกของคางที่น้อยลง[11]
- รูปร่างของกะโหลกศีรษะอย่างอื่น ๆ ที่เปลี่ยนไปเช่นกะโหลกมีลักษณะที่กลมขึ้น และมีโครงจมูกที่เล็กลง[10]
- การเกิดขึ้นของ styloid process ของกระดูกขมับ (เป็นกระดูกรูปร่างแหลมยื่นออกมาจากกะโหลกศีรษะใต้หูเพียงเล็กน้อย)
- การเกิดขึ้นของกล่องเสียงที่เคลื่อนตำแหน่งลงเมื่อโตขึ้น คือในเด็กวัยต้น อยู่ที่ระดับ C1-C3 ของกระดูกสันหลัง และจะเคลื่อนลงจากตำแหน่งนั้นเมื่อเจริญเติบโตขึ้น[109][110]
- การเจริญเติบโตของส่วนต่าง ๆ ในร่างกายที่เร็วช้าและมีลำดับที่ไม่เหมือนกัน และระยะเวลาที่นานกว่าที่จะเติบโตเป็นผู้ใหญ่[11]
ความเปลี่ยนแปลงทางพฤติกรรม
แก้แม้ว่า จะมีความแตกต่างทางพฤติกรรมที่สำคัญหลายอย่างระหว่างเอปกับมนุษย์ เช่นความสามารถในการใช้ภาษา แต่นักวิชาการทั้งหลายก็ยังไม่มีข้อยุติว่า จะสามารถใช้อะไรเป็นเครื่องแสดงความแตกต่างทางพฤติกรรมเหล่านี้ในบรรดาหลักฐานทางซากดึกดำบรรพ์ที่สามารถเห็นได้[11]
วิวัฒนาการก่อน โฮโม
แก้วิวัฒนาการของไพรเมต
แก้
ประวัติวิวัฒนาการของไพรเมตโดยซากดึกดำบรรพ์ย้อนเวลาไปประมาณ 55 ล้านปีก่อนในช่วงที่มีอากาศร้อน[5] สปีชีส์ของไพรเมตที่เก่าที่สุด (ที่รู้จัก) ก็คือสัตว์คล้ายมาโมเสทสกุล Teilhardina ที่อยู่ในทวีปยุโรป อเมริกาเหนือ และเอเชีย ระหว่าง 47 ถึง 56 ล้านปีก่อน ในต้นสมัย Eocene (ซึ่งอยู่ในกลางยุคพาลีโอจีน)[111] แต่ว่า หลักฐานโดยใช้ molecular clock (นาฬิกาอาศัยโครงสร้างโมเลกุล) ในปี ค.ศ. 2009 บอกเป็นนัยว่า ไพรเมตอาจเป็นสัตว์ที่เก่าแก่ยิ่งกว่านั้น คือเริ่มขึ้นในกลางยุคครีเทเชียสประมาณ 85 ล้านปีก่อน[112] ทันสมัยช่วงพวกไดโนเสาร์ซึ่งไปยุติที่ท้ายยุคครีเทเชียสที่ 66 ล้านปีก่อน และหลักฐานทางกายวิภาคและทางบรรพชีวินวิทยาอื่นในปี ค.ศ. 2007 แสดงว่า เป็นสัตว์ใกล้ชิดที่สุดกับกับสัตว์อันดับบ่าง[10]

แม้ว่า จะยังไม่มีหลักฐานทางซากดึกดำบรรพ์ที่เก่ากว่า 55.8 ล้านปี หลักฐานทางดีเอ็นเอบอกว่า มีการแยกสายพันธุ์ของไพรเมตก่อนหน้านั้นคือ[A]
- บรรพบุรุษของลีเมอร์แห่งเกาะมาดากัสการ์ ที่ 63 ล้านปี
- บรรพบุรุษของทาร์เซียร์แห่งเกาะในเอเชียอาคเนย์ ที่ 58 ล้านปี
- บรรพบุรุษของลิงโลกใหม่ (สาย Platyrrhini) ที่ 44 ล้านปี
นักวิชาการ[113] ได้สรุปว่า ไพรเมตในยุคต้น ๆ เกิดการขยายพันธุ์ไปทั่วทวีปยูเรเชีย และสายพันธุ์ที่สืบไปถึงเอปแอฟริการวมทั้งมนุษย์ ได้อพยพจากยุโรปและเอเชียตะวันตกไปทางใต้เข้าไปยังทวีปแอฟริกา ไพรเมตในเขตร้อนที่มีชีวิตรอดมาได้จนถึงปลายสมัย Eocene ซึ่งเห็นได้มากที่สุดในซากดึกดำบรรพ์ในชั้นหินยุคพาลีโอจีนปลายสมัย Eocene และต้นสมัยโอลิโกซีน ที่แอ่งใกล้เมือง Faiyum ทางทิศตะวันตกเฉียงใต้จากกรุงไคโร ประเทศอียิปต์ เป็นต้นตระกูลไพรเมตเขตร้อนที่ยังมีเหลือในปัจจุบันทั้งหมด เช่น
- ลีเมอร์ของเกาะมาดากัสการ์ (Lemuroidea)
- ลอริสของเอเชียอาคเนย์ (Lorisoidea)
- สัตว์วงศ์ Galagidae (อังกฤษ: galago, bush babies) ของแอฟริกา
- ลิงใน infraorder "Simiiformes" รวมทั้งลิงใน parvorder "Platyrrhini" หรือลิงโลกใหม่ (New World monkey), และ parvorder "Catarrhini" หรือวงศ์ลิงโลกเก่า (Old World monkey คือวงศ์ Cercopithecidae) บวกวงศ์ของเอป ซึ่งรวมมนุษย์ด้วย (ดูหัวข้อการจัดชั้นและการใช้ชื่อในบทความ)
สมัยโอลิโกซีน
แก้ในบรรดาซากดึกดำบรรพ์ที่พบใกล้เมือง Faiyum มีการค้นพบ
- สกุล Parapithecus เป็นไพรเมตที่เกิดขึ้นก่อนการแยกสายพันธุ์ของ "Platyrrhini" และ "Catarrhini" เป็นไพรเมตที่เป็นญาติกับบรรพบุรุษของเอป (รวมทั้งมนุษย์) อาจเป็นต้นตระกูลลิงโลกเก่า มีชีวิตในช่วงต้นสมัย Ogliocene ปลายสมัย Eocene[114]
- "เอป" สกุล Propliopithecus มีชีวิตอยู่กลางสมัยโอลิโกซีน อยู่ในวงศ์ Pliopithecidae ซึ่งเป็นวงศ์ของเอปที่เก่าแก่ที่สุดที่พบในซากดึกดำบรรพ์ อาจเป็นสกุลเดียวกับ หรือสืบสายมาจากสกุล Aegyptopithecus มีฟันกรามที่คล้ายกับเอป[114] เชื่อว่าเป็นสัตว์บรรพบุรุษหรือว่าเป็นญาติใกล้ชิดกับบรรพบุรุษมนุษย์[R]
- สกุล Aegyptopithecus เป็น "Catarrhini" ยุคต้น ๆ ที่เกิดขึ้นก่อนการแยกสายพันธุ์ของลิงโลกเก่าและเอป (รวมทั้งมนุษย์) อาจเป็นสกุลเดียวกับ Propliopithecus มีชีวิตระหว่าง 33 ถึง 35 ล้านปีก่อน มีลักษณะคล้ายเอปยิ่งกว่า Propliopithecus[114] เชื่อว่าเป็นสัตว์บรรพบุรุษหรือว่าเป็นญาติใกล้ชิดกับบรรพบุรุษมนุษย์[R]
ในปี ค.ศ. 2010 มีการค้นพบ Catarrhini สกุล Saadanius ในซาอุดีอาระเบีย มีอายุประมาณ 28 ถึง 29 ล้านปีก่อน (สมัยโอลิโกซีน กลาง) ที่ผู้ค้นพบเสนอว่าเป็นญาติกับบรรพบุรุษร่วมกันสุดท้ายของลิงโลกเก่าและเอป (รวมทั้งมนุษย์)[116] แม้ว่าจะยังไม่ชัดเจนว่า สปีชีส์ไหนเป็นต้นตระกูลของเอป แต่หลักฐานทางดีเอ็นเอแสดงว่า มีการแยกสายพันธุ์ของลิงโลกเก่าและเอปออกที่ 28 ล้านปี[A] ช่วงระยะปลายสมัยโอลิโกซีน กับต้นสมัยไมโอซีน
สมัยไมโอซีน
แก้ในสมัยไมโอซีน ตอนต้น ประมาณ 22 ล้านปีก่อน การมีลิง Catarrhini รุ่นต้น ๆ มากมายหลายประเภทที่ได้ปรับตัวอยู่บนต้นไม้ในแอฟริกาตะวันออก บอกเป็นนัยว่า มีประวัติยาวนานก่อนหน้านั้นของการเกิดความหลากหลายของสปีชีส์ต่าง ๆ ในสาย Catarrhini ซากดึกดำบรรพ์จากสมัยไมโอซีน ตอนกลาง (12 ถึง 16 ล้านปีก่อน) มีส่วนของลิงสกุล Victoriapithecus ซึ่งเป็นลิงโลกเก่าที่เก่าแก่ที่สุด (ไม่ใช่สายมนุษย์)[117] ส่วนเอปไร้หาง (ที่เป็นสายมนุษย์) เก่าแก่ที่สุดที่พบเป็นของสกุล Proconsul[S] มีอายุประมาณ 16 ถึง 25 ล้านปีก่อน ซึ่งเป็นช่วงท้ายสมัยโอลิโกซีน ต้นสมัยไมโอซีน อยู่ในแอฟริกาตะวันออก เป็นสัตว์ที่ลักษณะหลายอย่างคล้ายกับเอปปัจจุบันเช่นไม่มีหาง[119]
ส่วนสกุลอื่น ๆ ที่เชื่อกันว่าอยู่ในสายพันธุ์ของเอป (รวมทั้งมนุษย์) ที่พบจนกระทั่งถึง 13 ล้านปีก่อน (สมัยไมโอซีน กลาง) รวมสกุล Rangwapithecus, Dendropithecus, Limnopithecus, Nyanzapithecus, Afropithecus (17.5 ล้านปีในแอฟริกาและซาอุดิอาระเบีย), Griphopithecus (16.5 ล้านปีในเอเชียตะวันตก), Equatorius (10-16 ล้านปี), Heliopithecus (16 ล้านปีจากซาอุดิอาระเบีย), Kenyapithecus (15 ล้านปี), Nacholapithecus (15 ล้านปี), และ Dryopithecus (12 ล้านปีในยุโรป) โดยมากมาจากแอฟริกาตะวันออก
.JPG)
เริ่มตั้งแต่สมัยไมโอซีน กลางคือประมาณ 16 ถึง 17 ล้านปีก่อน ก็เริ่มพบเอปเป็นพวกแรก ๆ นอกแอฟริกา เพราะว่าในช่วงเวลานี้ เกิดทางเชื่อมกันระหว่างแอฟริกากับยูเรเชีย และเอปได้อพยพออกนอกแอฟริกาผ่านป่าฝนที่เกิดขึ้นต่อเนื่องเป็นป่าเดียวกัน ในขณะที่มีสัตว์จากยูเรเชียก็ได้อพยพเข้าไปในแอฟริกาด้วย[119] หลักฐานทางอณูชีววิทยาแสดงว่า สายพันธุ์ของชะนี (วงศ์ Hylobatidae) แยกสายพันธุ์ออกจากสายวงศ์ลิงใหญ่ ในช่วง 17 ล้านปีก่อน[A] แม้ว่าจะยังไม่ปรากฏซากดึกดำบรรพ์ที่แสดงหลักฐานทางบรรพบุรุษของพวกชะนี ซึ่งอาจมีกำเนิดจากกลุ่มเอปที่ยังไม่เป็นที่รู้จักในเอเชียอาคเนย์
การมีสัตว์วงศ์ลิงใหญ่ (บางสกุลจัดเข้าวงศ์ย่อย Homininae ด้วย) ที่อาจเป็นบรรพบุรุษของมนุษย์ หรือเป็นญาติของบรรพบุรุษมนุษย์ ในกลางสมัยไมโอซีน ในที่ไกล ๆ กัน เช่นสกุล Otavipithecus[T] จากถ้ำในประเทศนามิเบีย ที่แอฟริกาใต้, และ Pierolapithecus (มีชีวิตในสมัยไมโอซีน)[113] กับ Dryopithecus (9 ถึง 12 ล้านปีก่อน)[115] จากประเทศฝรั่งเศส สเปน และออสเตรีย ในยุโรป เป็นหลักฐานของความหลายหลากของวงศ์ลิงใหญ่ในแอฟริกาและบริเวณเมดิเตอร์เรเนียน ในสมัยที่มีอากาศค่อนข้างอุ่นและสม่ำเสมอในช่วงสมัยไมโอซีน ตอนต้นและตอนกลาง ลิงใหญ่ที่เกิดใหม่สุด (ที่พบซากดึกดำบรรพ์) ในสมัยไมโอซีน ก็คือสกุล Oreopithecus ซึ่งมาจากชั้นถ่านหินในประเทศอิตาลีมีอายุประมาณ 7 ถึง 9 ล้านปีก่อน[119]
จากบรรดาสปีชีส์ของวงศ์ลิงใหญ่เหล่านี้ นักวิทยาศาสตร์ยังไม่มีข้อมูลพอที่จะกำหนดได้ว่า พวกไหนเป็นสปีชีส์บรรพบุรุษของลิงใหญ่และมนุษย์ที่ยังมีชีวิตอยู่ในปัจจุบัน[13] แม้ว่าจะมีการเสนอว่าน่าจะเป็นสกุล Griphopithecus (14 ถึง 16.5 ล้านปีก่อน) จากเยอรมนีและตุรกี[113]
การแยกสายพันธุ์ของวงศ์ลิงใหญ่ (Miocene กลาง)
แก้- โดยหลักฐานทางอณูชีววิทยา
- ลิงอุรังอุตัง (สกุล Pongo) แยกออกจากสายพันธุ์ Homininae ประมาณ 14 ล้านปีก่อน (สมัยไมโอซีน กลาง)[A]
- สัตว์วงศ์ย่อย Homininae ที่ยังไม่สูญพันธุ์คือลิงโบโนโบ ลิงชิมแปนซี (ทั้งสองในสกุล Pan) และลิงกอริลลา (สกุล Gorilla) ซึ่งเป็นสัตว์ที่ใกล้กับมนุษย์ที่สุด[11] โดยที่จีโนมของมนุษย์และลิงชิมแปนซีมีความคล้ายคลึงกันที่ระดับประมาณ 95-99%[11][45][44] ซึ่งเป็นความคล้ายคลึงกันที่แสดงถึงความมีสายพันธุ์เป็นพี่น้องกัน (sister taxon) หรือแม้แต่อยู่ในสกุลเดียวกัน[15]
- ลิงกอริลลา (สกุล Gorilla) แยกออกจากสายพันธุ์มนุษย์ที่ประมาณ 6 ถึง 10 ล้านปีก่อน[10]
- ลิงชิมแปนซีและลิงโบโนโบ (สกุล Pan) แยกออกจากสายพันธุ์มนุษย์ที่ประมาณ 4 ถึง 8 ล้านปีก่อนซึ่งอยู่ในช่วงปลายสมัยไมโอซีน (ซึ่งเป็นส่วนปลายของยุคนีโอจีน)[10]
- โดยหลักฐานซากดึกดำบรรพ์
- บรรพบุรุษของลิงอุรังอุตัง (proto-orangutan) สามารถใช้สกุล Sivapithecus (7 ถึง 12.5 ล้านปีก่อน) จากประเทศอินเดียเป็นตัวแทนได้[113]
- ส่วนสปีชีส์ที่ใกล้กับบรรพบุรุษร่วมกันสุดท้ายของลิงกอริลลา ลิงชิมแปนซี และมนุษย์ สามารถใช้สกุล Nakalipithecus (9.8 ถึง 9.9 ล้านปีก่อน) พบในประเทศเคนยา[121] และ Ouranopithecus (7.4-9.6 ล้านปี) พบในประเทศกรีซ[122] เป็นตัวแทนได้
ส่วนซากดึกดำบรรพ์ของบรรพบุรุษลิงกอริลลาและบรรพบุรุษลิงชิมแปนซีนั้นมีจำกัด ทั้งการสงวนสภาพที่ไม่ดี (เพราะดินของป่าดิบชื้นที่เป็นที่อยู่ของลิงมักจะมีสภาพเป็นกรดซึ่งทำลายกระดูก) และความเอนเอียงในการคัดตัวอย่าง (sampling bias) เพื่อจะเลือกหาซากที่เป็นของมนุษย์ อาจก่อให้เกิดปัญหานี้ ส่วน Homininae ประเภทอื่น ๆ (รวมทั้งสายพันธุ์มนุษย์) น่าจะเกิดการปรับตัวให้เข้ากับสิ่งแวดล้อมที่แห้งกว่านอกเขตศูนย์สูตร พร้อม ๆ กับสัตว์เลี้ยงลูกด้วยนมอื่น ๆ เช่นแอนทิโลป ไฮยีน่า สุนัข หมู ช้าง และม้า เมื่อถึงประมาณ 8 ล้านปีก่อน เพราะเขตศูนย์สูตรได้เกิดการหดตัวลง
มีหลักฐานทางซากดึกดำบรรพ์น้อยเกี่ยวกับการแยกสายพันธุ์ของเผ่า hominini[B] จากลิงกอริลลาและลิงชิมแปนซี แต่ว่า การเดินด้วยสองเท้า (bipedalism) ซึ่งเป็นการปรับตัวพื้นฐานที่สุดของสัตว์เผ่า Hominini ซึ่งเป็นสายพันธุ์ของมนุษย์ที่ลิงชิมแปนซีได้แยกออกไปแล้ว ก็เริ่มปรากฏในสัตว์สองเท้าแรกสุดสปีชีส์ Sahelanthropus tchadensis[7] มีอายุ 7 ล้านปีก่อน (ปลายสมัยไมโอซีน)[13] หรือ Orrorin tugenensis[8] มีอายุ 6.2 ล้านปีก่อน[13] โดยมีสกุล Ardipithecus ซึ่งเป็นสัตว์สองเท้าที่มีหลักฐานชัดเจนกว่า[9] มีอายุ 5.8 ล้านปีก่อน[13] ตามมาทีหลัง[U]
สปีชีส์เหล่านี้ล้วนแต่มีการอ้างว่า เป็นบรรพบุรุษเดินด้วยสองเท้าของ hominin ที่เกิดต่อ ๆ มา แต่ว่าทุกกรณีก็มีนักวิชาการบางพวกที่ยังไม่เห็นด้วย[123][75][124] ดังนั้น จึงเป็นไปได้ว่า สปีชีส์หนึ่งหรือหลายสปีชีส์เหล่านี้เป็นสัตว์บรรพบุรุษของสายพันธุ์ลิงใหญ่แอฟริกา (คือลิงชิมแปนซีหรือลิงกอริลลา) คืออาจจะเป็นบรรพบุรุษที่ hominin มีร่วมกับลิงใหญ่แอฟริกา หรืออาจจะมี Sahelanthropus เป็นบรรพบุรุษสุดท้ายร่วมกันระหว่างชิมแปนซีและมนุษย์[7] ปัญหาเกี่ยวกับความสัมพันธ์ของซากดึกดำบรรพ์เหล่านี้กับสายพันธุ์มนุษย์ยังไม่มีข้อยุติ
hominin ก่อน Homo
แก้
จากสปีชีส์ในยุคต้น ๆ ที่กล่าวเป็นต้นแล้วนั้น hominin สกุล Australopithecus (เผ่าย่อย Australopithecina) ก็เกิดวิวัฒนาการในแอฟริกาตะวันออกประมาณ 3.9 ล้านปีก่อน ต้นสมัยไพลโอซีน[13] ก่อนที่จะแพร่พันธุ์ไปทั่วทวีป และมีซากดึกดำบรรพ์ที่พบครั้งสุดท้ายที่ 1.2 ล้านปีก่อน[63] ในระยะเวลาช่วงนั้น กลุ่มสปีชีส์ที่เรียกว่า gracile australopithecine เป็นกลุ่มที่เกิดก่อน มีหลายสปีชีส์รวมทั้ง A. anamensis (4.2 ล้านปี), A. afarensis (3.9 ล้านปี[126]), A. africanus (3.03 ล้านปี), และ A. sediba (1.98 ล้านปี) แต่ว่า นักวิชาการไม่มีมติร่วมกันว่า กลุ่มสปีชีส์ที่เรียกว่า robust australopithecine ที่เกิดต่อ ๆ มา รวมทั้ง A. aethiopicus (2.7 ล้านปี), A. boisei (2.3 ล้านปี), และ A. robustus (2 ล้านปี) ควรจะจัดเป็นสมาชิกของสกุล Australopithecus หรือไม่ ถ้าเป็น ทั้งสามสปีชีส์ก็จะมีทวินามดังที่กล่าวแล้ว แต่ว่า ถ้าควรจะอยู่ในอีกสกุลหนึ่ง ก็จะมีสกุลเป็นของตนเอง คือสกุล Paranthropus รวมทั้งหมดแล้ว เผ่าย่อย Australopithecina มี
- สกุล Australopithecus (1.7-4.2 ล้านปี กลุ่ม gracile australopithecine) รวมสปีชีส์ A. anamensis, A. afarensis, A. africanus, A. bahrelghazali, A. garhi และ A. sediba
- สกุล Kenyanthropus (3.2-3.5 ล้านปี) รวมสปีชีส์ Kenyanthropus platyops (บางที่รวมเข้ากับ Australopithecus)
- สกุล Paranthropus (1.2-2.7 ล้านปี กลุ่ม robust australopithecine) รวมสปีชีส์ P. aethiopicus, P. boisei และ P. robustus
สปีชีส์ที่มีหลักฐานทางซากดึกดำบรรพ์มากที่สุดก็คือ A. afarensis คือมีการพบซากเป็นหลายร้อย พบในที่ต่าง ๆ รวมทั้งประเทศเอธิโอเปียเหนือ (รวมทั้งซากของลูซี่) ประเทศเคนยา และประเทศแอฟริกาใต้ ส่วนซากของ robust australopithecine เช่นสปีชีส์ P. robustus และ P. boisei โดยเฉพาะ มีอยู่มากในที่ต่าง ๆ ในแอฟริกาใต้รวมทั้งเขตสงวน Kromdraai และเขตมรดกแห่งชาติ Swartkrans ในประเทศแอฟริกาใต้, และรอบ ๆ ทะเลสาบ Turkana ในประเทศเคนยา
ในบรรดาสกุลเหล่านี้ อาจจะมีสปีชีส์หนึ่งจากกลุ่ม gracile australopithecine คือ A. garhi[127] หรือ A. sediba[128] หรือ A. afarensis[15] หรือ A. africanus[129][V] หรือสปีชีส์อื่นที่ยังไม่พบ ที่กลายมาเป็นต้นตระกูลของสกุล โฮโม
วิวัฒนาการของสกุล โฮโม
แก้สมาชิกเก่าแก่ที่สุดของสกุล Homo ก็คือ H. habilis ซึ่งเกิดวิวัฒนาการที่ 2.3 ล้านปีก่อน[12][C] เป็นสปีชีส์แรกที่มีหลักฐานชัดเจนว่าใช้เครื่องมือหิน[D] ที่สร้างโดยมีระดับความซับซ้อนที่เรียกว่าเทคโนโลยีหิน Oldowan เป็นชื่อตามสถานที่คือโกรกธาร Olduvai gorge ที่พบตัวอย่างเครื่องมือหินเป็นครั้งแรก นักวิทยาศาสตร์บางพวกพิจารณา Homo rudolfensis ที่ค้นพบต่อมา ซึ่งมีร่างกายใหญ่กว่า แต่มีสัณฐานเหมือนกับซากของ H. habilis ว่าเป็นสปีชีส์ต่างหาก แต่บางพวกก็พิจารณาว่า เป็นพวกเดียวกับ H. habilis คือ มีลักษณะที่แตกต่างกันบ้างภายในสปีชีส์เดียวกัน หรืออาจจะเป็นความแตกต่างระหว่างเพศเลยด้วยซ้ำ (คือเป็นของเพศชาย)[15]
วิวัฒนาการทางกายภาพที่ชัดเจนที่สุดของมนุษย์สกุล Homo ที่แตกต่างจากสายพันธุ์ Australopithecina ก็คือ ขนาดกะโหลกศีรษะที่ใหญ่ขึ้น คือจากประมาณ 450 ซม3 ใน Australopithecus garhi[127] มาเป็น 610 ซม3 ใน H. habilis[12] ในช่วงล้านปีต่อมา กระบวนการขยายขนาดสมองก็ดำเนินต่อไป คือ ภายในสกุล Homo เอง ขนาดกะโหลกศีรษะได้เพิ่มขึ้นเป็นสองเท่าจาก H. habilis ผ่าน H. erectus/ergaster ไปยัง H. heidelbergensis ที่ 1,250 ซม3 เมื่อ 600,000 ปีก่อน[131] หลักฐานซากดึกดำบรรพ์ของมนุษย์ต่อมาคือสปีชีส์ H. ergaster/erectus ซึ่งเกิดขึ้นประมาณ 1.9 ล้านปีก่อน[24] เป็นมนุษย์สกุล Homo พวกแรกที่อพยพออกไปตั้งถิ่นฐานทั่วทวีปแอฟริกา ทวีปเอเชีย และทวีปยุโรป เริ่มตั้งแต่ 1.8 ล้านปีก่อน[26] แต่ก็มีกลุ่มหนึ่งของ H. erectus ที่ดำรงอยู่ในทวีปแอฟริกา (หรือเรียกว่า African H. erectus) ซึ่งบางครั้งจัดเป็นสปีชีส์ต่างหากคือ H. ergaster (ซึ่งได้รับการเสนอว่า เป็นบรรพบุรุษของ H. sapiens)[132] เชื่อกันว่านี้เป็นสปีชีส์แรก ๆ ที่สามารถควบคุมไฟ[25]
ซากดึกดำบรรพ์ช่วงเปลี่ยนสภาพ (transitional fossil) ที่เก่าที่สุดระหว่าง H. ergaster/erectus และมนุษย์ที่เกิดต่อ ๆ มา คือกลุ่มมนุษย์โบราณ มาจากทวีปแอฟริกา แต่ซากที่เก่าแก่ที่สุดนอกแอฟริกาได้พบที่โบราณสถาน Dmanisi ประเทศจอร์เจีย เขตคอเคซัส (H. georgicus แม้ว่านักวิทยาศาสตร์โดยมากจัดให้อยู่ในกลุ่ม H. erectus/ergaster) มีอายุประมาณ 1.8 ล้านปี[133]
ต่อมาเริ่มตั้งแต่ประมาณ 1.2 ล้านปีก่อน สปีชีส์ลูกหลานมนุษย์ H. ergaster/erectus ที่เรียกว่า มนุษย์โบราณ (Archaic humans) จึงปรากฏว่าได้ตั้งถิ่นฐานแล้วทั้งในทวีปแอฟริกาและทั่วทวีปยูเรเชีย[40] คือวิวัฒนาการเป็นมนุษย์สปีชีส์ (อายุในวงเล็บแสดงซากเก่าแก่ที่สุดที่พบ)
- H. antecessor (1.2 ล้านปี พบในยุโรป[134]),
- H. heidelbergensis (600,000 ปี พบในแอฟริกาและยุโรป[131]),
- H. rhodesiensis (500,000-600,000 ปี[27] ปี พบในแอฟริกา) และ
- H. neanderthalensis (200,000-300,000 ปี พบในยุโรปและเอเชียตะวันตก[104])
ส่วนซากดึกดำบรรพ์ที่เก่าแก่ที่สุดของมนุษย์ที่มีกายวิภาคเหมือนมนุษย์ปัจจุบันมาจากยุคหินกลาง (แอฟริกา) ประมาณ 315,000 ปีก่อน จากโบราณสถาน Jebel Irhoud ประเทศโมร็อกโก[G] โดยยังมีซากเก่าแก่อื่น ๆ รวมทั้งกลุ่มซากดึกดำบรรพ์ Omo remains จากประเทศเอธิโอเปีย โดยเก่าที่สุดมีอายุ 195,000 ปี[82], ของสปีชีส์ย่อย Homo sapiens idaltu จากหมู่บ้าน Herto ในโบราณสถาน Herto Formation ในประเทศเอธิโอเปีย โดยมีอายุเกือบ 160,000 ปี[81], และจากถ้ำ Skhul ในประเทศอิสราเอลที่มีอายุประมาณ 90,000-100,000 ปี[82] ซึ่งเป็นซากของมนุษย์ปัจจุบันที่เก่าแก่ที่สุดนอกแอฟริกา[31]
ตามทฤษฎีกำเนิดมนุษย์ปัจจุบันเร็ว ๆ นี้จากแอฟริกา การอพยพที่เป็นต้นเหตุของประชากรในโลกปัจจุบันทั้งหมด เริ่มขึ้นที่ประมาณ 70,000 ปีก่อน แล้วมนุษย์ปัจจุบันก็ขยายถิ่นฐานไปทั่วโลก แทนที่ hominin รุ่นก่อน ๆ ถ้าไม่โดยการแข่งขันกัน ก็โดยการผสมพันธุ์กัน และได้เข้าไปตั้งถิ่นฐานในทวีปยูเรเชียและในเขตโอเชียเนียก่อน 40,000 ปีก่อน และในทวีปอเมริกาทั้งเหนือใต้ก่อน 14,500 ปีก่อน[135] (ดูรายละเอียดเพิ่มเติมที่การอพยพย้ายถิ่นฐานของมนุษย์)


H. sapiens เป็นสปีชีส์เดียวที่ยังเหลือในสกุล โฮโม แม้ว่า สปีชีส์อื่น ๆ ของ โฮโม ที่สูญพันธุ์แล้วอาจจะเป็นบรรพบุรุษของ H. sapiens แต่หลายสปีชีส์ก็น่าจะเป็น "ลูกพี่ลูกน้อง" ของเรามากกว่า เพราะได้แตกสาขาไปจากบรรพบุรุษของเรา ยังไม่มีมติที่เห็นพ้องกันว่า กลุ่มไหนควรจะเป็นสปีชีส์ต่างหาก กลุ่มไหนควรจะนับในสปีชีส์ย่อย (subspecies) ความไม่ลงเอยกันในบางกรณีเป็นเพราะความขาดแคลนหลักฐานทางซากดึกดำบรรพ์ และในกรณีอื่น เป็นเพราะวิธีการจัดสปีชีส์[138] ทฤษฎีปัมพ์สะฮารา (Sahara pump theory) ซึ่งแสดงว่าทะเลทรายสะฮาราบางครั้งมีแหล่งน้ำเพียงพอที่จะข้ามได้ เสนอคำอธิบายที่เป็นไปได้อย่างหนึ่งเกี่ยวกับความต่าง ๆ กันของสปีชีส์ในสกุล โฮโม[139] แม้บทความนี้จะไม่มีรายละเอียด
ตามทฤษฎีมหันตภัยภูเขาไฟทะเลสาบโตบา (Toba catastrophe theory) ที่นักมานุษยวิทยาและนักโบราณคดีบางส่วนเห็นด้วย ซูเปอร์ภูเขาไฟของทะเลสาบโตบาบนเกาะสุมาตราในประเทศอินโดนีเซียเกิดระเบิดเมื่อประมาณ 70,000 ปีก่อนโดยมีผลไปทั่วโลก ทำให้มนุษย์โดยมากในช่วงนั้นเสียชีวิตและสร้างคอคอดประชากรที่มีผลต่อสายพันธุ์ที่สืบกันมาจนถึงทุกวันนี้[140]
H. rudolfensis
แก้H. rudolfensis เป็นชื่อที่ให้กับมนุษย์ที่มีอายุ 2.4 ถึง 1.8 ล้านปีก่อน ที่มีตัวอย่างต้นแบบที่ค้นพบในปี ค.ศ. 1972 ในประเทศเคนยาใกล้ทะเลสาบ Lake Turkana โดยทีมของริชาร์ด ลีกคี การค้นพบชิ้นส่วนขากรรไกรของสปีชีส์นี้ ในปี ค.ศ. 2012 โดยทีมของมีฟ ลีกคี เชื่อว่าเป็นหลักฐานสนับสนุนอย่างชัดเจนว่า เป็นสปีชีส์ต่างหาก เพราะเป็นสปีชีส์ที่มีกะโหลกศีรษะ โดยเฉพาะในส่วนกระดูกขากรรไกรที่แตกต่างจาก H. habilis อย่างเห็นได้ชัด ซึ่งให้หลักฐานว่าเป็นสกุล Homo ยุคต้น ๆ ที่อยู่ร่วมกับ H. habilis[141]
แต่ว่า เรื่องนี้ยังไม่มีข้อยุติ เพราะมีนักวิชาการบางท่านที่เสนอว่า อาจจะเป็นตัวอย่างของสกุล Australopithecus บางพวกก็เสนอว่า ควรย้ายเข้าไปรวมในสกุล Australopithecus บางท่านก็เสนอว่าเป็นซากของ H. habilis แต่ยังไม่มีหลักฐานยืนยัน และบางท่านก็เสนอว่า ตัวอย่างที่ได้ยังน้อยเกินที่จะยุติเรื่องนี้ นอกจากนั้นแล้ว ก็ยังไม่ชัดเจนว่า เป็น H. Habiis หรือ H. rudolfensis หรือสปีชีส์อื่นที่ยังไม่พบ ที่เป็นต้นตระกูลของมนุษย์ปัจจุบัน[141][11]
สปีชีส์นี้มีขนาดกะโหลกศีรษะเฉลี่ยที่ 750 ซม3 ซึ่งใหญ่กว่าของ H. habilis ที่เกิดทีหลัง แต่มีลักษณะหลายอย่างรวมทั้งฟันกรามที่ใหญ่ ที่เหมือนกับสายพันธุ์ Australopithecine มากกว่า[141]
H. habilis และ H. gautengensis
แก้Homo habilis มีชีวิตอยู่ในครึ่งแรกของสมัยไพลสโตซีนประมาณ 1.5 ถึง 2.3 ล้านปีก่อน[C] โดยอาจวิวัฒนาการมาจาก australopithecine ตัวอย่างซากดึกดำบรรพ์ที่เป็นแบบก็คือซากศพหมายเลข OH 7 ที่พบในปี 2503 (ค.ศ. 1960) ณ โบราณสถาน Olduvai Gorge ในประเทศแทนซาเนีย ต่อมาในปี 2507 จึงได้จัดเป็นสปีชีส์ต่างหากโดยตั้งชื่อเป็น H. habilis (แปลว่า มือคล่องแคล่ว หรือชำนาญ อังกฤษ: handy man) เพราะว่า ซากดึกดำบรรพ์มักจะพบพร้อมกับเครื่องมือหินแบบ Oldowan และเชื่อว่า มนุษย์พวกนี้สามารถแปลงหินธรรมชาติให้เป็นเครื่องมือหินได้ เป็นมนุษย์สกุล Homo ที่รูปร่างสัณฐานคล้ายกับมนุษย์ปัจจุบันน้อยที่สุด (คือลักษณะบางอย่างคล้ายกับ australopithecine มากกว่า) โดยยกเว้นสปีชีส์ที่มีปัญหาจัดเข้าในสกุลมนุษย์เช่นกันคือ H. rudolfensis ตั้งแต่นั้นมา การจัดอยู่ในสกุลก็ได้สร้างข้อถกเถียงกันอย่างไม่มีที่ยุติ[12] ขนาดที่เล็กและลักษณะล้าหลังทำให้ผู้ชำนาญการ (รวมทั้ง ริชาร์ด ลีกคีเอง) เสนอว่าควรกัน H. habilis ออกจากสกุล Homo แล้วใส่ไว้ใน Australopithecus โดยจัดเป็น Australopithecus habilis[142]
นักบรรพมานุษยวิทยาชาวอังกฤษ-เคนยา หลุยส์ ลีกคี เป็นบุคคลแรกที่เสนอว่ามีมนุษย์จำพวกนี้ โดยภรรยาคือ แมรี ลีกคี เป็นผู้พบฟันสองซี่แรกของ H. habilis ในปี 2498 ซึ่งต่อมาระบุว่าเป็น ฟันน้ำนม ซึ่งพิจารณาว่าเป็นตัวระบุชนิดสัตว์ได้ยากโดยไม่เหมือนกับฟันแท้ ต่อมาในปี 2502 แมรีจึงได้ค้นพบกะโหลกศีรษะของเด็กชายที่มีสมองเล็ก ใบหน้าใหญ่ ฟันเขี้ยวเล็ก และฟันเคี้ยวขนาดใหญ่ ทำให้สปีชีส์นี้ได้ชื่อเล่นอีกอย่างว่า นายกะเทาะเปลือกถั่ว (อังกฤษ: The Nutcracker man)[143]
H. habilis เชื่อว่าเป็นผู้ใช้เทคโนโลยีเครื่องมือหินแบบ Olduwan ในยุคหินเก่าต้น เพื่อฆ่าและแล่หนังสัตว์[144] เป็นเครื่องมือที่ล้ำหน้ากว่าเครื่องมืออื่น ๆ ที่เคยใช้มาทั้งหมด ทำให้ได้เปรียบในการใช้ชีวิตในสิ่งแวดล้อมที่ก่อนหน้านี้ยากเกินสำหรับสัตว์อันดับวานร แต่ H. habilis จะเป็นสายพันธุ์มนุษย์แรกที่สามารถใช้เครื่องมือหินหรือไม่ ก็ยังไม่ชัดเจน เพราะว่าแม้แต่ Australopithecus garhi ซึ่งมีอายุประมาณ 2.6 ล้านปีก่อน ก็ยังพบพร้อมกับเครื่องมือหินด้วย[D]
H. habilis เตี้ย โดยผู้ชายสูงเฉลี่ย 1.3 เมตร มีแขนยาวเหมือนกับ australopithecine ที่เชื่อว่าเป็นบรรพบุรุษ และจะดูไม่สมส่วนเทียบกับมนุษย์ปัจจุบัน แต่ว่า ก็ยังมีใบหน้าที่ยื่นออกน้อยกว่า australopithecine มีฟันและสันคิ้วที่เล็กกว่า australopithecine แต่เมื่อเทียบกับมนุษย์ปัจจุบันแล้ว ฟันจะจัดเป็นแนวกลมเหมือนกัน แม้ฟันเขี้ยวจะค่อนข้างใหญ่ มีใบหน้าและสันคิ้วที่หนากว่า[12] แม้ว่าจะมีรูปร่างสัณฐานที่คล้ายเอป แต่ซากของ H. habilis บ่อยครั้งก็อยู่ร่วมกับเครื่องมือที่ทำจากหินแบบง่าย ๆ[D][12] และอาจจะกระดูกสัตว์[W]
ขนาดสมองเฉลี่ยของมนุษย์กลุ่มนี้อยูที่ 610 ซม3[12] ซึ่งใหญ่กว่าของ australopithecine ประมาณ 50% โดยเฉลี่ย แต่ก็ยังเล็กกว่าสมองมนุษย์ปัจจุบันที่ 1,330 ซม³ พอสมควร[12] ส่วนงานสร้างใหม่เสมือนที่ตีพิมพ์ในปี 2558 ประมาณปริมาตรภายในกะโหลกที่ระหว่าง 729-824 ซม3 ซึ่งใหญ่กว่าขนาดที่เคยรายงานมาทั้งหมด[148]
ผู้ชำนาญการโดยมากสมมุติว่า ทั้งเชาว์ปัญญาและการจัดระเบียบทางสังคมของมนุษย์กลุ่มนี้ ซับซ้อนยิ่งกว่าที่พบโดยทั่วไปใน australopithecine หรือในลิงชิมแปนซี H. habilis ใช้เครื่องมือโดยหลักเพื่อหากินซากสัตว์ เช่น ตัดเนื้อออกจากซากสัตว์ ไม่ใช่เพื่อป้องกันตัวหรือล่าสัตว์ อย่างไรก็ดี แม้ว่าจะใช้เครื่องมือ มนุษย์กลุ่มนี้ก็ยังไม่ชำนาญการล่าสัตว์เหมือนกับมนุษย์กลุ่มอื่น ๆ เพราะหลักฐานดึกดำบรรพ์จำนวนมากแสดงว่า มนุษย์กลุ่มนี้เป็นอาหารของสัตว์ล่าเหยื่อขนาดใหญ่ เช่น เสือเขี้ยวดาบสกุล Dinofelis ซึ่งมีขนาดพอ ๆ กับ เสือจากัวร์ (หนัก 120 กก. สูง 70 ซม.)[149]
ขนาดข้อต่อของแขนขาของมนุษย์จำพวกนี้คล้ายกับ A. afarensis ซึ่งไม่คล้ายของมนุษย์ปัจจุบัน ซึ่งแสดงนัยว่า บางที H. sapiens อาจไม่คล้ายกับมนุษย์พวกนี้จริง ๆ ตามที่เสนอ แต่สัดส่วนร่างกายของมนุษย์พวกนี้ก็สมกับหลักฐานกะโหลกศีรษะและฟันที่แสดงว่า สัมพันธ์อย่างใกล้ชิดกับ H. erectus[150]
H. habilis มักพิจารณาว่าเป็นบรรพบุรุษของ H. ergaster ที่ผอมงามกว่า (gracile) และฉลาดซับซ้อนมากกว่า ซึ่งก็เป็นบรรพบุรุษของสปีชีส์ที่เหมือนมนุษย์ยิ่งขึ้น คือ H. erectus ข้อถกเถียงว่า ซากดึกดำบรรพ์ที่พบแล้วทั้งหมดได้จัดเข้ากับสปีชีส์นี้อย่างถูกต้องหรือไม่ ก็ยังดำเนินต่อไป โดยมีนักบรรพมานุษยวิทยาที่มองหน่วยอนุกรมวิธานนี้ว่า เป็นโมฆะ เพราะว่า ประกอบด้วยตัวอย่างซากดึกดำบรรพ์ของสัตว์ทั้งสกุล Australopithecus และ Homo[151]
มนุษย์กลุ่มนี้อยู่กับไพรเมตคล้ายมนุษย์อื่น ๆ รวมทั้ง Paranthropus boisei ที่บางกลุ่มอยู่ได้เป็นพัน ๆ ปี แต่ว่า โดยอาจเป็นเพราะนวัตกรรมทางเทคโนโลยีและอาหารที่เฉพาะเจาะจงน้อยกว่า H. habilis ได้เป็นบรรพบุรุษของมนุษย์สปีชีส์ต่าง ๆ จำนวนหนึ่ง เทียบกับ P. boisei และ robust australopithecine ที่หายไปจากบันทึกซากดึกดำบรรพ์ และ H. habilis ยังอาจอยู่ร่วมกัน H. erectus ในแอฟริกาเป็นเวลากว่า 500,000 ปี[152]
มนุษย์นี้ตอนแรกพิจารณาว่าเป็นสปีชีส์แรกในสกุล Homo จนกระทั่งงานวิเคราะห์โครงกระดูกที่พบในอดีตได้ตีพิมพ์ในเดือนพฤษภาคม 2553 แล้วเสนอสปีชีส์ใหม่ คือ H. gautengensis จากประเทศแอฟริกาใต้ โดยผู้เสนอเชื่อว่าเป็นสปีชีส์เก่าแก่ที่สุดในสกุล Homo[153] ส่วนงานปี 2556 พบส่วนหนึ่งของกระดูกขากรรไกรที่มีอายุราว 2.8 ล้านปีก่อน โดยมีหมายเลข LD 350-1 พบในโบราณสถาน Ledi-Geraru ในบริเวณอฟาร์ของเอธิโอเปีย[154] โดยซากดึกดำบรรพ์ที่พบพิจารณาว่า เป็นหลักฐานเก่าแก่ที่สุดของสกุล Homo ที่เคยพบจนถึงปีนั้น และดูเหมือนจะเป็นซากดึกดำบรรพ์ในระหว่าง Australopithecus และ H. habilis เป็นมนุษย์ที่มีชีวิตหลังจากการเปลี่ยนแปลงสภาพอากาศแบบฉับพลัน ที่สิ่งแวดล้อมแบบป่าและทางน้ำ ได้กลายเป็นทุ่งหญ้าสะวันนาอย่างรวดเร็ว[155]
H. ergaster/erectus
แก้ในสมัยไพลสโตซีนช่วงต้น คือ 1.5 ถึง 2 ล้านปีก่อน ในทวีปแอฟริกา H. erectus เกิดการวิวัฒนาการให้มีสมองใหญ่ขึ้นแล้วใช้เครื่องมือหินที่ซับซ้อนยิ่งขึ้นและสามารถควบคุมไฟได้ ความแตกต่างเช่นนี้และอย่างอื่น ๆ เพียงพอที่จะให้นักมานุษยวิทยาจัดเป็นสปีชีส์ใหม่ คือ H. erectus[156] ซากดึกดำบรรพ์แรกของ H. erectus มีการค้นพบโดยนายแพทย์ชาวดัตช์ ยูจีน ดูบัวส์ ในปี ค.ศ. 1891 ที่เกาะชวาในประเทศอินโดนีเซีย ซึ่งตอนแรกเขาตั้งชื่อว่า Pithecanthropus erectus โดยอาศัยสัณฐานที่พิจารณาว่าเป็นสปีชีส์ในระหว่างมนุษย์และเอป[157] ตัวอย่างซากดึกดำบรรพ์ที่มีชื่อเสียงของ H. erectus ก็คือมนุษย์ปักกิ่ง[158] และ "Turkana boy" (หรือ "Nariokotome Boy")[133] ส่วนตัวอย่างอื่น ๆ มีการพบในเอเชีย (โดยเฉพาะประเทศอินโดนีเซีย) แอฟริกา และยุโรป
H. erectus นั้นมีชีวิตอยู่ในระหว่าง 27,000-1.9 ล้านปีก่อน[X][24] และดังนั้น ถ้าหลักฐานที่พบทั้งหมดสามารถจัดเป็นสปีชีส์เดียวกันได้จริง ๆ ก็จะเป็นสกุล Homo ที่ดำรงอยู่ได้กว่า 1.5 ล้านปี[13] ซึ่งนานกว่ามนุษย์สกุล Homo อื่นทั้งหมด เชื่อกันว่า H. erectus สืบสายพันธุ์มาจากสกุลก่อน ๆ เช่น Ardipithecus หรือ Australopithecus หรือจาก Homo สปีชีส์อื่น ๆ เช่น H. habilis หรือ H. ergaster แต่ว่าทั้ง H. erectus, H. ergaster, และ H. habilis ก็ล้วนแต่มีช่วงอายุที่คาบเกี่ยวกัน ดังนั้น จึงอาจจะเป็นสายพันธุ์ต่างหาก ๆ ที่สืบมาจากบรรพบุรุษเดียวกัน[159]
สมองของ H. erectus มีขนาดประมาณ 725-1,250 ซม3[11] H. erectus ในยุคต้น ๆ ดูเหมือนจะสืบทอดเทคโนโลยีเครื่องมือหิน Oldowan มาจากมนุษย์ยุคก่อน ๆ แล้วพัฒนาให้เป็นเทคโนโลยีหินแบบ Acheulean เริ่มตั้งแต่ 1.76 ล้านปีก่อน[158]
ส่วนหลักฐานถึงการควบคุมไฟได้ของ H. erectus ตั้งต้นแต่ 400,000 ปีก่อนได้รับการยอมรับจากนักวิชาการโดยมาก และหลักฐานที่เก่ากว่านั้นก็เริ่มที่จะได้รับการยอมรับจากนักวิทยาศาสตร์ โดยมีหลักฐานที่อ้างการใช้ไฟที่เก่าที่สุดมาจากแอฟริกาใต้ พบในปี ค.ศ. 2011 ที่ 1.8 ล้านปีก่อน[25] และมีหลักฐานของเครื่องมือหินเผาไฟที่ได้รับการยอมรับเพิ่มขึ้นเรื่อย ๆ พบในปี ค.ศ. 2004 ในประเทศอิสราเอลโดยมีอายุ 790,000 ปี[160] ส่วนการใช้ไฟเพื่อหุงอาหารนั้นยังเป็นเรื่องที่ยังไม่มีข้อยุติ[25][161][162][163] เพราะยังไม่มีซากดึกดำบรรพ์ที่แสดงหลักฐานการหุงอาหารอย่างชัดเจน ส่วนพวกที่เห็นว่ามีการใช้ไฟเพื่อหุงอาหารเสนอว่า การหุงอาหารเป็นการช่วยปล่อยสารอาหารและทำให้ย่อยได้ง่าย และช่วยทำลายพิษในพืชบางประเภท[164] นอกจากนั้นแล้ว H. erectus ยังเสนอว่า เป็นมนุษย์พวกแรกที่ใช้แพข้ามทะเล เพราะพบเครื่องมือหินบนเกาะ Flores ในอินโดนีเซียที่ไม่ปรากฏทางไปทางบก[31]
H. erectus เป็นมนุษย์สกุล Homo พวกแรกที่อพยพออกไปตั้งถิ่นฐานทั่วทวีปแอฟริกา ทวีปเอเชีย และทวีปยุโรป เริ่มตั้งแต่ 1.8 ล้านปีก่อน[26] แต่มีกลุ่มหนึ่งของ H. erectus ที่ดำรงอยู่ในทวีปแอฟริกา (หรือเรียกว่า African H. erectus) ซึ่งบางครั้งจัดเป็นสปีชีส์ต่างหากคือ H. ergaster ซึ่งต่อมาได้รับการเสนอว่า เป็นบรรพบุรุษของ H. sapiens[132] นักบรรพมานุษยวิทยาบางพวกปัจจุบันใช้นามว่า H. ergaster สำหรับมนุษย์สปีชีส์นี้ที่พบบางส่วนในแอฟริกา (ทั้งหมดจากประเทศเคนยาใกล้ทะเลสาบ Lake Turkana) ที่มีโครงสร้างกระดูกและฟันที่ต่างจาก H. erectus เพียงเล็กน้อย และใช้นามว่า H. erectus สำหรับซากดึกดำบรรพ์พบในที่ ๆ เหลือรวมทั้งแอฟริกาด้วย[11]
ส่วน H. georgicus จากประเทศจอร์เจีย ซึ่งตอนแรกเสนอว่าเป็นสปีชีส์ในระหว่าง H. habilis และ H. erectus[165] เดี๋ยวนี้ได้จัดเป็นสปีชีส์ย่อยของ H. erectus[166] คือ H. erectus georgicus
H. antecessor และ H. cepranensis
แก้ซากดึกดำบรรพ์เหล่านี้ได้รับการเสนอว่าเป็นสปีชีส์ในระหว่าง H. erectus และ H. heidelbergensis
- H. antecessor มาจากซากดึกดำบรรพ์พบในสเปนและอาจจะในอังกฤษ[167] มีอายุระหว่าง 8 แสนปี-1.2 ล้านปี ในระหว่าง ค.ศ. 1994-1996 มีการค้นพบซากดึกดำบรรพ์ของมนุษย์ 80 คนที่อาจจะเป็นของสปีชีส์นี้ในโบราณสถานในประเทศสเปน มีตัวอย่างของกระดูกมนุษย์ที่ปรากฏว่ามีการแล่เนื้อออก ซึ่งอาจจะบอกว่าสปีชีส์นี้กินเนื้อมนุษย์ (เพราะไม่มีหลักฐานว่า มนุษย์พวกนี้มีการทำพิธีกรรมเกี่ยวกับศพ) จากชิ้นส่วนซากดึกดำบรรพ์ที่มีอยู่น้อยมาก มีการประมาณว่า H. antecessor มีความสูงที่ 160-180 ซม และผู้ชายหนักประมาน 90 กก มีสมองขนาดเฉลี่ยประมาณ 1,000 ซม3 ซึ่งเล็กกว่าขนาดเฉลี่ยมนุษย์ปัจจุบัน และน่าจะมีร่างกายที่แข็งแรงกว่า H. heidelbergensis สปีชีส์นี้เป็นหลักฐานของมนุษย์สกุล Homo ที่เก่าแก่ที่สุดในยุโรป มีผู้เสนอว่า สปีชีส์นี้เป็นบรรพบุรุษของทั้ง Neanderthal และมนุษย์ปัจจุบันและเป็นสปีชีส์ที่เกิดในทวีปแอฟริกาแล้วอพยพออกมาทางยุโรป[134]
- H. cepranensis มีซากดึกดำบรรพ์เป็นยอดของกะโหลกศีรษะ (skull cap) เดียวที่พบในอิตาลี ในปี ค.ศ. 1994 ประเมินว่ามีอายุประมาณ 700,000-1 ล้านปี เป็นซากดึกดำบรรพ์ของมนุษย์ที่เก่าแก่ที่สุดซากหนึ่งในยุโรป[168] โดยทั่ว ๆ ไปแล้ว นักวิทยาศาสตร์จัดซากดึกดำบรรพ์นี้ว่าเป็นซากเก่าที่สุดของ H. heidelbergensis หรือเป็นสปีชีส์ที่เป็นบรรพบุรุษต้นกำเนิดของทั้งมนุษย์ปัจจุบันและ H. neanderthalensis[169]
H. heidelbergensis/rhodesiensis
แก้H. heidelbergensis หรือบางครั้งเรียกว่า H. rhodesiensis[134] เป็นมนุษย์ที่สูญพันธุ์ไปแล้ว ที่มีชีวิตอยู่ในแอฟริกา ยุโรป และเอเชียประมาณระหว่าง 250,000-600,000 ปีก่อน[11] แต่อาจเก่าแก่ถึง 1.4 ล้านปีก่อน[170] เป็นมนุษย์กลุ่มแรกที่มีสมองขนาดใกล้กับมนุษย์ แต่มีร่างกายที่แข็งแรงกว่า[11] มีการเสนอว่าสืบเชื้อสายมาจาก H. ergaster/erectus ในแอฟริกา และเป็นบรรพบุรุษโดยตรงของ H. sapiens ในแอฟริกา, ของ H. neanderthalensis ในยุโรป, และของมนุษย์กลุ่ม Denisovan ในเอเชียกลาง[136][Y]
ทั้ง H. antecessor และ H. heidelbergensis น่าจะสืบเชื้อสายมาจาก H. ergaster ในแอฟริกาเพราะว่า มีสัณฐานคล้ายกันมาก แต่เพราะว่า H. heidelbergensis มีกะโหลกหุ้มสมองที่ใหญ่กว่ามาก คือปกติประมาณ 1,200 ซม3 (เทียบกับ H. ergaster/erectus ที่ 850 ซม3 และ H. antecessor ที่ 1,000 ซม3) และมีเครื่องมือและพฤติกรรมที่ทันสมัยกว่า จึงได้รับการจัดให้อยู่ในสปีชีส์ต่างหาก ผู้ชายสูงโดยเฉลี่ยประมาณ 175 ซม หนัก 62 กก และผู้หญิงสูงโดยเฉลี่ยประมาณ 158 ซม หนัก 55 กก[173] เป็นสปีชีส์ที่โดยเฉลี่ยสูงกว่ามนุษย์สาย Neanderthal เพียงเล็กน้อย[174] แต่ก็มีนักบรรพมานุษยวิทยาบางท่านที่อ้างว่า มีกลุ่ม "ยักษ์" กลุ่มหนึ่งที่ปกติสูงกว่า 213 ซม อยู่ในแอฟริกาใต้ประมาณ 300,000-500,000 ปีก่อน[175]
แม้ว่า H. heidelbergensis จะเกิดวิวัฒนาการขึ้นในแอฟริกา แต่ก็ได้อพยพออกไปตั้งถิ่นฐานทั่วยุโรปและอาจจะในเอเชีย (จีนและอิสราเอล)[173] โดย 500,000 ปีก่อน เครื่องมือหินที่ใช้ในตอนต้นเป็นเทคโนโลยี Acheulean เหมือนกับที่ H. ergaster/erectus ใช้ แต่ในกาลต่อ ๆ มาบางที่ในยุโรป จึงพบเครื่องมือที่ซับซ้อนยิ่งขึ้นทำจากเขากวาง กระดูก และไม้ โดยทำเป็นมีดแล่ เครื่องทุบ หอกไม้ และหอกหินมีด้ามไม้ นอกจากนั้นแล้ว H. heidelbergensis ยังร่วมมือกันล่าสัตว์ใหญ่อาจจะรวมทั้งแรดและฮิปโปโปเตมัสเป็นต้น[131] H. rhodesiensis (อังกฤษ: Rhodesian Man) มีชีวิตอยู่ช่วง 125,000-400,000 ปีก่อน มีขนาดสมองประมาณ 1,100-1,230 ซม3[176] นักวิจัยปัจจุบันโดยมากจัด H. rhodesiensis ไว้ในกลุ่มของ H. heidelbergensis[11] นักวิจัยบางท่านให้ความเห็นว่า เป็นไปได้ที่ Rhodesian Man จะเป็นบรรพบุรุษของ Homo sapiens idaltu (เป็นชื่อของสปีชีส์ย่อยของมนุษย์ปัจจุบันที่มีซากดึกดำบรรพ์ที่เก่าแก่ที่สุดซากหนึ่งค้นพบในแอฟริกา) ซึ่งก็เป็นบรรพบุรุษของ Homo sapiens sapiens ซึ่งก็คือมนุษย์ปัจจุบัน[81]
นีแอนเดอร์ทาลและ Denisovan
แก้H. neanderthalensis (อังกฤษ: Neanderthal) มีชื่อภาษาอังกฤษตั้งขึ้นตามชื่อหุบเขาที่พบซากดึกดำบรรพ์เป็นครั้งแรก (คือหุบเขา Neander ในประเทศเยอรมัน) ใช้ชีวิตอยู่ในยุโรปและเอเชียประมาณ 28,000-300,000 ปีก่อน โดยมีขนาดสมองเฉลี่ยที่ 1,500 ซม3[104] ซึ่งใหญ่กว่าขนาดเฉลี่ยในมนุษย์ปัจจุบันที่ 1,3303[103][Z] งานวิจัยปี ค.ศ. 2008 โดยใช้แบบจำลองคอมพิวเตอร์พบว่า ทารกของมนุษย์ Neanderthal เมื่อคลอดจะมีสมองขนาดเท่ากับของทารกมนุษย์ปัจจุบัน แต่สมองจะใหญ่กว่าเมื่อถึงวัยผู้ใหญ่[180]
มีหลักฐาน (ค.ศ. 1997, 2004, 2008) โดยการหาลำดับดีเอ็นเอของไมโทคอนเดรีย (mitochondrial DNA ตัวย่อ mtDNA) ที่แสดงว่า ไม่มีการแลกเปลี่ยนยีนโดยเป็นนัยสำคัญคือไม่มีการผสมพันธุ์กันระหว่าง H. neanderthalensis และ H. sapiens ดังนั้น สองกลุ่มนี้จึงเป็นสปีชีส์ที่แยกจากกันโดยมีบรรพบุรุษเดียวกันเมื่อประมาณ 500,000-600,000 ปีก่อน[104] โดยอาจมีบรรพบุรุษเป็น H. heidelbergensis/rhodesiensis[136] แต่ว่า งานหาลำดับดีเอ็นเอทั้งจีโนมของมนุษย์กลุ่มนี้ในปี ค.ศ. 2010 กลับแสดงว่า มีการผสมพันธุ์กับมนุษย์ปัจจุบันเมื่อประมาณ 45,000-80,000 ปีก่อน (ประมาณช่วงเวลาที่มนุษย์ปัจจุบันออกจากแอฟริกา แต่ก่อนที่จะไปตั้งถิ่นฐานในยุโรป เอเชีย และที่อื่น ๆ) มนุษย์ปัจจุบันที่ไม่ใช่คนแอฟริกาเกือบทั้งหมดมีดีเอ็นเอ 1-4% สืบมาจากมนุษย์กลุ่มนี้[181] ซึ่งเข้ากับงานวิจัยเร็ว ๆ นี้ที่แสดงว่า การแยกออกจากกันของอัลลีลในมนุษย์บางพวกเริ่มขึ้นที่ 1 ล้านปีก่อน แต่ว่า การตีความหมายข้อมูลจากงานวิจัยทั้งสองที่แสดงผลแตกต่างกันนี้ ยังมีประเด็นที่น่าสงสัย[182][183]
นีแอนเดอร์ทาลคล้ายมนุษย์ปัจจุบันแต่ลักษณะใบหน้าบางอย่างก็ไม่เหมือน และมีลำตัวที่กำยำล่ำสันที่เหมาะกับการอยู่ใน่ที่หนาวมากกว่า หน้ามีสันคิ้ว หน้าผากค่อนข้างเรียบที่เทลาดไปด้านหลัง มีส่วนจมูกที่ยื่นออก มีหลุมตาใหญ่และกลม มีจมูกใหญ่ ทั่วไปจะเตี้ยกว่ามนุษย์ปัจจุบันคือผู้ชายสูงประมาณ 168 ซม และผู้หญิงประมาณ 156 ซม แม้จะมีกระดูกและกล้ามเนื้อที่แข็งแรงกว่า มีฟันใหญ่ และมีขากรรไกรที่แข็งแรง กว่ามนุษย์ปัจจุบัน[104] นีแอนเดอร์ทาลแข็งแรงกว่ามนุษย์ปัจจุบันมาก โดยเฉพาะที่แขนและมือ[184]
นีแอนเดอร์ทาลใช้เครื่องมือหินก้าวหน้าที่เรียกว่าเทคโนโลยี Mousterian ซึ่ง H. sapiens ต้น ๆ ก็ใช้เหมือนกัน หลังจากนั้นในช่วงที่มนุษย์ปัจจุบันเริ่มเข้าไปสู่ทวีปยุโรป ก็เริ่มผลิตเครื่องมือโดยวิธีใหม่ที่เรียกว่า Chatelperronian ซึ่งให้ผลได้ใบมีดที่ H. sapiens ใช้เหมือนกัน ดังนั้นจึงเกิดสันนิษฐานว่า อาจลอกแบบกัน หรือติดต่อค้าขายกัน[104]
นีแอนเดอร์ทาลรู้จักสร้างเตาและสามารถควบคุมไฟ อาศัยหนังสัตว์ห่อหุ้มร่างกาย[AA] แม้อาจจะยังไม่รู้จักเย็บ[104] แต่ก็เจาะรูแล้วผูกเข้าด้วยกัน มีพืชสำหรับทานน้อยกว่ามนุษย์ในแอฟริกาโดยเฉพาะในหน้าหนาว จึงต้องล่าสัตว์ต่าง ๆ ทานโดยใช้หอกรวมทั้ง กวางเรนเดียร์และกวางแดง และพวกที่อยู่ตามชายทะเลอาจทานหอยมอลลัสกา แมวน้ำ โลมา และปลา เป็นอาหารด้วย นีแอนเดอร์ทาลใช้เครื่องประดับและมีพิธีฝังผู้ตาย รวมการฝังเครื่องบูชาศพเช่นดอกไม้[AB] เป็นมนุษย์รุ่นแรก ๆ ที่มีพฤติกรรมแบบสัญลักษณ์เยี่ยงนี้ที่ไม่พบในมนุษย์ก่อน ๆ[68]
- ดูรายละเอียดอื่น ๆ เกี่ยวกับดีเอ็นเอของมนุษย์โบราณที่หัวข้อการอพยพย้ายถิ่นฐานของมนุษย์
H. neanderthalensis และ H. sapiens อาจจะอยู่ร่วมกันในยุโรปเป็นเวลานานถึง 10,000 ปี ซึ่งเป็นช่วงเวลาที่ประชากรมนุษย์ปัจจุบันเกิดการประทุมากขึ้นเป็นสิบเท่าทำให้มีจำนวนมากกว่า Neanderthal อย่างมาก ซึ่งอาจทำให้ Neanderthal แข่งขันสู้ไม่ได้เพียงเพราะเหตุแห่งจำนวนนั้น[185] แล้วนำไปสู่การสูญพันธุ์เมื่อประมาณ 30,000 ปีก่อน นอกจากสันนิษฐานนี้แล้ว ก็ยังมีสันนิษฐานอื่น ๆ อีกหลายข้อเกี่ยวกับการสูญพันธุ์เป็นต้นว่า
- มนุษย์ปัจจุบันมีเทคโนโลยีและพฤติกรรมที่ก้าวหน้ากว่า ทำให้ Neanderthal แข่งขันสู้ไม่ได้[104] (ดูรายละเอียดที่หัวข้อ ยุคหินเก่ากลางและปลาย)
- ความเปลี่ยนแปลงของภูมิอากาศ[104]
ในปี ค.ศ. 2008 นักโบราณคดีที่ทำงานที่ถ้ำ Denisova ในเทือกเขาอัลไตของเขตไซบีเรียได้ค้นพบกระดูกชิ้นเล็กชิ้นหนึ่งจากนิ้วก้อยของเด็กสายพันธุ์ Denisovan[104] และสิ่งประดิษฐ์ต่าง ๆ รวมทั้งกำไลมืออันหนึ่งที่ขุดได้จากชั้นหินในระดับเดียวกันโดยหาอายุได้ประมาณ 40,000 ปี นอกจากนั้นแล้ว เนื่องจากว่ามีดีเอ็นเอที่รอดอยู่ในซากหินเนื่องจากถ้ำมีอากาศเย็น จึงมีการหาลำดับดีเอ็นเอของทั้ง mtDNA และของทั้งจีโนม ในขณะที่การแยกสายพันธุ์พบใน mtDNA ย้อนไปไกลกว่าที่คาดคิด ลำดับของจีโนมของนิวเคลียสเซลล์กลับบอกเป็นนัยว่า Denisovan อยู่ในสายพันธุ์เดียวกับ Neanderthal โดยมีการแยกสายพันธุ์เป็นสองสปีชีส์หลังจากบรรพบุรุษที่ได้แยกสายพันธุ์ออกจากของมนุษย์ปัจจุบันก่อนหน้านั้น[Y]
เป็นที่รู้กันอยู่แล้วว่า มนุษย์ปัจจุบันอาจอยู่ร่วมกับ Neanderthal ในยุโรปเป็นช่วงเวลากว่า 10,000 ปี และการค้นพบนี้แสดงความเป็นไปได้ว่า มนุษย์ปัจจุบัน, Neanderthal, และ Denisovan อาจมีช่วงเวลาที่อยู่ร่วมกัน การมีอยู่ของสาขาต่าง ๆ ของมนุษย์เช่นนี้ อาจทำให้ภาพพจน์เกี่ยวกับมนุษย์ในสมัยไพลสโตซีนช่วงปลายซับซ้อนขึ้นกว่าที่คิด นอกจากนั้นแล้ว ยังมีหลักฐานที่แสดงว่าจีโนมของชาวเมลานีเซียปัจจุบันประมาณ 6% สืบมาจาก Denisovan ซึ่งแสดงถึงการผสมพันธุ์ในระดับจำกัดระหว่างมนุษย์ปัจจุบันกับ Denisovan ในเอเชียอาคเนย์[32]
มีรายละเอียดทางกายวิภาคของ Denisovan น้อย เพราะซากที่ได้พบมาทั้งหมดมีแต่กระดูกนิ้วมือ, ฟันสองซี่ที่ได้หลักฐานทางดีเอ็นเอ, และกระดูกนิ้วเท้า นิ้วมือที่มีมีลักษณะกว้างและแข็งแรง เกินกว่าที่เห็นได้ในกลุ่มมนุษย์ปัจจุบัน ที่น่าแปลกใจก็คือ เป็นนิ้วของหญิง ซึ่งอาจจะแสดงว่า Denisovan มีร่างกายที่แข็งแรงมาก อาจจะคล้ายพวกมนุษย์ Neanderthal[172]
H. floresiensis
แก้
H. floresiensis ซึ่งมีชีวิตอยู่ในช่วง 13,000-95,000 ปีก่อน มีชื่อเล่นว่า "ฮ็อบบิท" เพราะตัวเล็ก ซึ่งอาจเกิดขึ้นผ่านกระบวนการ insular dwarfism (ซึ่งสัตว์ใหญ่ย่อขนาดลงโดยผ่านหลายชั่วรุ่น เมื่อเกิดการจำกัดพื้นที่ เช่นย้ายไปอยู่บนเกาะ) อาจเคยมีชีวิตอยู่ร่วมกับมนุษย์ปัจจุบัน และมีนักวิชาการบางท่านที่เสนอว่า อาจมีความเกี่ยวข้องกับสัตว์ตำนานพื้นบ้าน (ของแหล่งที่ค้นพบสปีชีส์นี้) ที่เรียกว่า Ebu gogo เป็นสปีชีส์ที่น่าสนใจทั้งโดยขนาดและโดยความเก่าแก่ เพราะว่าเป็นตัวอย่างของสปีชีส์หลัง ๆ ของสกุล Homo ที่มีลักษณะสืบสายพันธุ์ (trait) ที่มนุษย์ปัจจุบันไม่มี กล่าวอีกนัยหนึ่งก็คือ แม้ว่ามนุษย์สปีชีส์นี้จะมีบรรพบุรุษร่วมกับมนุษย์ปัจจุบัน แต่ก็ได้เกิดการแยกสายพันธุ์ แล้วมีวิถีวิวัฒนาการเป็นของตนเอง โครงกระดูกหลักที่พบเชื่อว่าเป็นของหญิงอายุประมาณ 30 ปี พบในปี ค.ศ. 2003 บนเกาะ Flores ในประเทศอินโดนีเซีย โดยมีความเก่าแก่ประมาณ 18,000 ปี มีความสูงประมาณ 1 เมตร และมีขนาดสมองเพียงแค่ 380-420 ซม3[186] (ซึ่งเรียกว่าเล็กแม้ในลิงชิมแปนซี และมีขนาดเพียงแค่ 1/3 ของมนุษย์ปัจจุบัน)
อย่างไรก็ดี ก็ยังมีข้อถกเถียงกันว่า H. floresiensis ควรจะจัดเป็นสปีชีส์ต่างหากหรือไม่ เพราะนักวิทยาศาสตร์บางพวกเชื่อว่า เป็น H. sapiens ที่มีสภาพแคระโดยโรค[186] สมมติฐานนี้มีหลักฐานโดยส่วนหนึ่งว่า มนุษย์ปัจจุบันบางพวกที่เกาะ Flores ที่ค้นพบซากของสปีชีส์ เป็นคนพิกมี (pygmies ซึ่งมักหมายถึงกลุ่มชนที่ผู้ชายโดยเฉลี่ยมีความสูงต่ำกว่า 150-155 ซ.ม.) เพราะฉะนั้น คนพิกมีที่มีสภาพแคระโดยโรค อาจจะทำให้เกิดมนุษย์ที่มีรูปร่างคล้ายฮ็อบบิท ข้อขัดแย้งที่สำคัญอีกอย่างหนึ่งในเรื่องการจัดเป็นสปีชีส์ต่างหากก็คือ เครื่องมือที่พบเป็นชนิดที่ปกติค้นพบเฉพาะกับ H. sapiens[187]
ถึงกระนั้น สมมติฐานสภาพแคระโดยโรคก็ไม่สามารถอธิบายลักษณะอื่น ๆ ของสปีชีส์นี้ ซึ่งเหมือนกับลิงชิมแปนซี หรือกับ hominin ในยุคต้น ๆ เช่น Australopithecus และที่ไม่เหมือนกับมนุษย์ปัจจุบัน (ไม่ว่าจะมีหรือไม่มีโรค) ลักษณะที่ว่านี้ รวมลักษณะต่าง ๆ ของกะโหลกศีรษะ รูปร่างของกระดูกที่ข้อมือ ที่แขนท่อนปลาย ที่ไหล่ ที่เข่า และที่เท้า นอกจากนั้นแล้ว สมมติฐานยังไม่สามารถอธิบายการมีตัวอย่างหลายตัวอย่างที่มีลักษณะเหมือน ๆ กันเช่นนี้ ซึ่งบ่งว่าเป็นเรื่องสามัญในคนหมู่ใหญ่ ไม่ใช่เป็นลักษณะที่มีอยู่เฉพาะบุคคลเท่านั้น และมีงานวิจัยโดยคำนวณความคล้ายคลึงกันระหว่างสปีชีส์ (cladistic analysis) และโดยการวัดส่วนต่าง ๆ ของร่างกาย ที่สนับสนุนสมมติฐานว่าเป็นสปีชีส์ต่างหากจาก H. sapiens[186]
H. sapiens
แก้H. sapiens (คำวิเศษณ์ ว่า sapiens เป็นภาษาละตินแปลว่า ฉลาด[82]) มีชีวิตอยู่ตั้งแต่ประมาณ 300,000 ปีก่อน[G] จนถึงปัจจุบัน คือ ในระหว่าง 250,000-400,000 ปีก่อน เริ่มปรากฏแนวโน้มการขยายขนาดสมอง[E][103] และความซับซ้อนของเทคโนโลยีเครื่องมือหิน ในหมู่มนุษย์ ซึ่งเป็นหลักฐานของวิวัฒนาการที่เริ่มมาจาก H. ergaster/erectus (1.9 ล้านปีก่อน) จนถึง H. sapiens (300,000 ปีก่อน) และมีหลักฐานโดยตรงที่บอกว่า H. erectus อพยพออกจากแอฟริกาก่อน[26] แล้วจึงวิวัฒนาการเป็นสปีชีส์ H. sapiens โดยสืบมาจาก H. ergaster/erectus ที่ยังคงอยู่ในแอฟริกา[132] (ผ่าน "H. heidelbergensis/rhodesiensis"[136]) แล้วก็ได้อพยพต่อ ๆ ไปทั้งภายในและภายนอกแอฟริกาประมาณ 50,000-100,000 ปีก่อน ซึ่งในที่สุดก็ทดแทนมนุษย์สายพันธุ์ของ H. erectus/ergaster ในที่ต่าง ๆ ทั้งหมด[28] ทฤษฎีกำเนิดและการอพยพของบรรพบุรุษมนุษย์ปัจจุบันนี้มักจะเรียกว่า "Recent Single Origin" (ทฤษฎีกำเนิดเดียวเร็ว ๆ นี้), "Recent African Origin" (ทฤษฎีกำเนิดในแอฟริกาเร็ว ๆ นี้), หรือ "Out of Africa theory" (ทฤษฎีออกจากแอฟริกา) เป็นทฤษฏีที่ได้รับการยอมรับอย่างกว้างขวางที่สุดในหมู่นักวิทยาศาสตร์[78]
แต่ว่า หลักฐานที่มีไม่ได้ห้ามการผสมพันธุ์กัน (admixture[H]) ระหว่าง H. sapiens กับมนุษย์กลุ่ม Homo ก่อน ๆ และก็ยังไม่สามารถล้มทฤษฎีที่แข่งกันคือ Multiregional Origin (ทฤษฏีกำเนิดมนุษย์หลายเขตพร้อม ๆ กัน)[49] ซึ่งเป็นประเด็นขัดแย้งที่ยังไม่ยุติในสาขาบรรพมานุษยวิทยา (paleoanthropology) (ดูรายละเอียดเพิ่มขึ้นที่หัวข้อ "การอพยพย้ายถิ่นฐานของมนุษย์")
งานวิจัยปัจจุบันได้ทำให้ชัดเจนแล้วว่า มนุษย์ทั้งหมดมีกรรมพันธุ์ที่คล้ายกันในระดับสูง ซึ่งก็คือ ดีเอ็นเอของแต่ละคนคล้ายกันเกินกว่าที่ทั่วไปกับสปีชีส์อื่น ๆ (เช่นคนสองคนจากกลุ่มเดียวกันมักมีดีเอ็นเอที่คล้ายคลึงกันมากกว่าลิงชิมแปนซีจากกลุ่มเดียวกัน)[28] ซึ่งอาจจะเกิดเพราะเพิ่งเกิดวิวัฒนาการขึ้นเร็ว ๆ นี้ หรืออาจจะเป็นเพราะเกิดคอคอดประชากร ที่เป็นผลมาจากภัยธรรมชาติเช่น มหันตภัยภูเขาไฟทะเลสาบโตบา (Toba catastrophe)[32] ที่ทำให้ผู้คนในที่ต่าง ๆ ล้มตายเป็นจำนวนมาก ทำให้มนุษย์ปัจจุบันเป็นเชื้อสายสืบทอดมาจากบุคคลกลุ่มค่อนข้างเล็ก กล่าวอีกอย่างหนึ่งก็คือ มนุษย์ปัจจุบันมีลักษณะทางกรรมพันธุ์ที่ไม่เหมือนกันในจีโนมค่อนข้างน้อย เป็นเหมือนกับสถานการณ์ที่ชนกลุ่มเล็ก ๆ อพยพเข้าไปอยู่ในสิ่งแวดล้อมใหม่ ๆ (จึงมีลูกหลานที่ต่างกันทางกรรมพันธุ์น้อย) และส่วนเล็กน้อยที่ไม่เหมือนกันนั้น ได้ปรากฏเป็นลักษณะต่าง ๆ เช่นสีผิว[188] และรูปร่างจมูก รวมทั้งลักษณะภายใน เช่น สมรรถภาพการหายใจในที่สูงอย่างมีประสิทธิภาพ[189][190]

ซากมนุษย์ H. sapiens/"ที่มีกายวิภาคปัจจุบัน" ที่เก่าแก่ที่สุด
แก้งานศึกษา 2 งานในวารสาร Nature เดือนกรกฎาคม 2560[191][192] กล่าวถึง
- การหาอายุของเคลือบฟันของฟันที่ได้พบก่อน (Irhoud 3) โดยวิธี electron spin resonance
- ซากดึกดำบรรพ์มนุษย์ที่ขุดพบใหม่
- การหาอายุของเครื่องมือหินแบบ Middle Stone Age (MSA) ในชั้นตะกอนเดียวกับซากดึกดำบรรพ์ใหม่โดยวิธี thermoluminescence
- การสำรวจพรรณสัตว์ในชั้นตะกอนเดียวกับซากดึกดำบรรพ์ใหม่ และการหาอายุของซากสัตว์
- การสร้างแบบจำลองกะโหลกศีรษะโดยใช้ซากดึกดำบรรพ์ใหม่
ที่สรุปว่า ได้ค้นพบบรรพบุรุษของมนุษย์ปัจจุบัน ซึ่งมีลักษณะทางกายวิภาคที่ยังสามารถจัดอยู่ในกลุ่ม H. sapiens[191][192][193][194][195][196][197][198] โดยเรียกว่าเป็น "H. sapiens ต้น ๆ"[194] หรือ "H. sapiens ที่กำลังดำเนินไปสู่การมีกายวิภาคปัจจุบัน"[198] และมีอายุประมาณที่ 300,000 ปีก่อน[G] หลักฐานซากดึกดำบรรพ์ที่ได้ มาจากทั้งที่มีอยู่ก่อนแล้วและการค้นพบใหม่ที่โบราณสถาน Jebel Irhoud ประเทศโมร็อกโก ผู้นำกลุ่มนักวิชาการสากลกลุ่มนี้ได้กล่าวว่า เป็นหลักฐานแสดงถึง[191][194][195]
- วิวัฒนาการของมนุษย์ปัจจุบันที่เป็นไปทั่วแอฟริกา (คือมีโบราณสถานที่ค้นพบมนุษย์เก่าแก่ทั้งทางเหนือ ทางตะวันออก และทางใต้) พร้อมกับเครื่องมือหินแบบ MSA โดยไม่จำกัดอยู่ทางด้านตะวันออกของทวีปดังที่นักวิชาการบางพวกอาจจะเคยคิดเท่านั้น และซากมนุษย์ในภาคเหนือ (โมร็อกโก) ก็ได้พบพร้อมกับซากดึกดำบรรพ์สัตว์ประเภทเดียวกับที่อยู่ทางแอฟริกาตะวันออกด้วย ซึ่งแสดงถึงการเชื่อมต่อกันระหว่างภูมิภาคทั้งสอง (ไม่ได้แยกจากกันโดยทะเลทรายสะฮาราเหมือนปัจจุบัน)
- วิวัฒนาการมาเป็นมนุษย์ปัจจุบันโดยเกิดที่ใบหน้าก่อน แล้วจึงตามด้วยกระดูกหุ้มสมองกับสมอง เพราะว่า แบบจำลองที่สร้างแสดงว่า มีใบหน้าคล้ายของมนุษย์ปัจจุบัน แต่กระดูกหุ้มสมองนั้นยาวกว่าและกลมน้อยกว่า โดยนักวิชาการท่านอื่น (Christopher Stringer) คาดว่า การเกิดใบหน้าแบบเรียบ ๆ อาจสัมพันธ์กับการพูดได้
แม้จะมีนักวิชาการอื่นบางพวกที่สนับสนุนแนวคิดเช่นนี้ แต่ก็มีผู้ที่ขัดแย้งหลัก ๆ ในสองประเด็นคือ
- เป็นการประเมินอายุกะโหลกศีรษะที่จำลองโดยปริยาย เพราะได้อายุจากฟันและเครื่องมือหินที่พบในหินชั้นเดียวกัน[194] และเป็นเทคนิคหาอายุที่ไม่เลือกใช้กับหลักฐานจากแอฟริกาตะวันออกที่ได้การยอมรับอย่างกว้างขวาง ดังนั้น อายุประเมินนี้ อาจต้องมีการทดสอบหรือทบทวนหลักฐานอื่น ๆ เพิ่มเติมในอนาคต[198]
- กะโหลกศีรษะที่จำลอง (โดยหลักคือหน้าผาก) ไม่เหมือนของมนุษย์ปัจจุบันพอ หรือว่า การจัดโครงสร้างกระดูกแบบต่าง ๆ กันเข้าในกลุ่มเดียวกัน ทำให้มองเห็นประวัติวิวัฒนาการของมนุษย์ได้ยาก จึงไม่ควรจัดเข้าในกลุ่มมนุษย์ H. sapiens[193]
ส่วนซากดึกดำบรรพ์ที่เก่าแก่ที่สุดของมนุษย์ "ที่มีกายวิภาคของมนุษย์ปัจจุบันต้น ๆ"[197] มาจากกลางยุคหินเก่า ประมาณ 200,000 ปีก่อน คือจากกลุ่มซากดึกดำบรรพ์ Omo remains จากประเทศเอธิโอเปีย โดยเก่าที่สุดมีอายุ 195,000 ปี[82] ส่วนซากอื่น ๆ รวมทั้ง
- สปีชีส์ย่อย Homo sapiens idaltu จากหมู่บ้าน Herto ในโบราณสถาน Herto Formation ในประเทศเอธิโอเปีย โดยมีอายุเกือบ 160,000 ปี มีขนาดสมองประมาณ 1,450 ซม3 และผู้ค้นพบให้ความเห็นว่า เป็นบรรพบุรุษของมนุษย์ปัจจุบัน[81][81]
- ซากจากถ้ำ Skhul ในประเทศอิสราเอลที่มีอายุประมาณ 90,000-100,000 ปี[82] ซึ่งเป็นซากของมนุษย์ปัจจุบันที่เก่าแก่ที่สุดนอกแอฟริกา[31]
การใช้เครื่องมือ
แก้_01_palmar_view.png)

ส่วนยื่น styloid process ที่สุดของกระดูกติดกับข้อมือ ของกระดูกฝ่ามือชิ้นที่สาม (ดูรูป) ช่วยให้กระดูกมือล็อกเข้ากับกระดูกข้อมือได้ ซึ่งทำให้นิ้วโป้งและนิ้วมืออื่น ๆ ที่ใช้จับวัตถุสิ่งของอยู่ สามารถสื่อแรงดันไปที่มือและข้อมือในระดับที่สูงขึ้นได้ ทำให้มนุษย์มีความคล่องแคล่วและมีกำลังในการทำและใช้เครื่องมือที่ซับซ้อน ลักษณะพิเศษทางกายนี้ทำมนุษย์ให้แตกต่างไปจากเอปและไพรเมตที่ไม่ใช่มนุษย์อื่น ๆ และไม่พบในซากมนุษย์ที่เก่ากว่า 1.8 ล้านปี[108] (คือไม่พบก่อนมนุษย์ Homo ergaster/erectus โดยประมาณ[90])
การใช้เครื่องมือได้รับการตีความหมายว่าเป็นนิมิตของสติปัญญา มีทฤษฎีว่า การใช้เครื่องมืออาจจะช่วยกระตุ้นวิวัฒนาการบางอย่างของมนุษย์ โดยเฉพาะในเรื่องการขยายขนาดของสมอง[E] ที่เป็นไปอย่างต่อเนื่อง แต่หลักทางบรรพชีวินวิทยายังไม่สามารถอธิบายเหตุของการขยายขนาดของสมองเป็นช่วงเวลาหลายล้านปีอย่างชัดเจน แม้ว่าจะเป็นอวัยวะที่ใช้พลังงานสูง คือ สมองมนุษย์ปัจจุบันกินพลังงานประมาณ 13 วัตต์ต่อวัน หรือ 260 กิโลแคลอรี ซึ่งเป็น 1/5 ของพลังงานที่ร่างกายใช้ทั้งหมดต่อวันโดยเฉลี่ย[199] แต่การใช้เครื่องมือเพิ่มขึ้นทำให้สามารถแล่เอาเนื้อสัตว์ซึ่งมีพลังงานสูงได้ และช่วยให้สามารถจัดการอาหารพืชที่มีพลังงานสูงได้ ดังนั้น นักวิจัยจึงได้เสนอว่า hominin ในยุคต้น ๆ อาจอยู่ใต้ความกดดันทางวิวัฒนาการเพื่อเพิ่มสมรรถภาพในการสร้างและใช้เครื่องมือ เพื่อที่จะลดขนาดทางเดินอาหารที่ต้องมีถ้าต้องบริโภคอาหารที่มีพลังงานต่ำ แล้วใช้พลังงานสำหรับบริหารทางเดินอาหารในการบริหารสมองแทน[200][201]
เวลาที่มนุษย์ยุคแรก ๆ เริ่มใช้เครื่องมือเป็นสิ่งที่กำหนดได้ยาก เพราะว่า เครื่องมือยิ่งง่าย ๆ เท่าไร (เช่น หินมีคม) ก็จะแยกแยะว่าเป็นวัตถุธรรมชาติหรือเป็นสิ่งประดิษฐ์ยากยิ่งขึ้นเท่านั้น มีหลักฐานบ้างว่า hominin สาย australopithecine (กำเนิด 4 ล้านปีก่อน) ใช้ก้อนหินแตกเป็นเครื่องมือ เมื่อ 3.4 ล้านปีก่อน แต่เรื่องนี้ยังไม่มีข้อยุติ[D]
ควรจะสังเกตว่า มีสัตว์หลายสปีชีส์ที่ทำและใช้เครื่องมือ[202] แต่เป็นสกุลต่าง ๆ ของมนุษย์ที่ทำและใช้เครื่องมือที่ซับซ้อนเหนือกว่าสัตว์อื่น ๆ ทั้งหมด เครื่องมือหินเก่าแก่ที่สุดที่ชัดเจนเป็นเครื่องมือหินเทคโนโลยี Oldowan จากประเทศเอธิโอเปีย อายุประมาณ 2.6 ล้านปี เป็นชื่อตามสถานที่คือโกรกธาร Olduvai gorge ที่พบตัวอย่างเครื่องมือหินเป็นครั้งแรก ซึ่งเกิดขึ้นก่อนสกุล Homo แรกสุด คือ H. habilis ที่ 2.3 ล้านปี และยังไม่มีหลักฐานว่ามีสกุล Homo ที่เกิดขึ้นโดย 2.5 ล้านปีก่อน แต่ว่า มีซากดึกดำบรรพ์ของ Homo ที่พบใกล้เครื่องมือ Oldowan แม้ว่าจะมีอายุประมาณเพียง 2.3 ล้านปี (คือเครื่องมือหินที่พบเก่าแก่กว่า) ซึ่งบอกเป็นนัยว่า บางทีมนุษย์ Homo อาจจะเป็นผู้สร้างและใช้เครื่องมือเหล่านี้ แต่เป็นเรื่องเป็นไปได้ที่ยังไม่มีหลักฐานที่ชัดเจน และมีนักวิชาการบางพวกที่สันนิษฐานว่าเป็น hominin สาย Australopithecine ที่สร้างเครื่องมือหินขึ้นเป็นพวกแรก[D]
นักวิชาการบางท่านให้ข้อสังเกตว่า hominin สกุล Paranthropus นั้นอยู่ร่วมกับสกุล Homo ในเขตที่พบเครื่องมือหิน Oldowan เป็นครั้งแรก ในช่วงเวลาเดียวกัน ถึงแม้ว่าจะไม่มีหลักฐานโดยตรงที่บ่งว่า Paranthropus เป็นผู้สร้างเครื่องมือ แต่ลักษณะทางกายวิภาคก็ให้หลักฐานโดยอ้อมถึงความสามารถในเรื่องนี้ แต่นักบรรพมานุษยวิทยาโดยมากมีมติพ้องกันว่า มนุษย์ Homo รุ่นต้น ๆ เป็นผู้สร้างและผู้ใช้เครื่องมือ Oldowan ที่พบโดยมาก ไม่ใช่ australopithecine เช่น Paranthropus คือ อ้างว่า เมื่อพบเครื่องมือ Oldowan กับซากมนุษย์ ก็จะพบ Homo ด้วย แต่บางครั้งจะไม่พบ Paranthropus[50]
ในปี ค.ศ. 1994 มีงานวิจัยที่ใช้กายวิภาคของหัวแม่มือที่สามารถจับสิ่งของร่วมกับนิ้วอื่นได้ (opposable thumb) เป็นฐานในการอ้างว่า ทั้งสกุล Homo และ Paranthropus สามารถสร้างและใช้เครื่องมือได้ คือได้เปรียบเทียบกระดูกและกล้ามเนื้อของหัวแม่มือระหว่างมนุษย์กับลิงชิมแปนซี แล้วพบว่า มนุษย์มีกล้ามเนื้อ 3 ชิ้นที่ชิมแปนซีไม่มี นอกจากนั้นแล้ว มนุษย์ยังมีกระดูกฝ่ามือ ที่หนากว่าและมีหัวกระดูกที่กว้างกว่า ซึ่งสามารถทำให้จับสิ่งของได้แม่นยำกว่าชิมแปนซี นักวิจัยยืนยันว่า กายวิภาคปัจจุบันของหัวแม่มือมนุษย์เป็นการตอบสนองทางวิวัฒนาการเกี่ยวกับการที่ต้องทำและใช้เครื่องมือ ซึ่ง hominii ทั้งสองสปีชีส์สามารถทำเครื่องมือได้[50]
เครื่องมือหิน
แก้-
!["หินคม" เป็นเครื่องมือหิน Oldowan ซึ่งพบครั้งแรกที่โกรกธาร Olduvai gorge เป็นเครื่องมือหินขั้นพื้นฐานที่สุดของมนุษย์ (ยุคหินเก่าต้น) เริ่มตั้งแต่ H. habilis[D]](//up.wiki.x.io/wikipedia/commons/thumb/d/de/Pierre_taill%C3%A9e_Melka_Kunture_%C3%89thiopie_fond.jpg/120px-Pierre_taill%C3%A9e_Melka_Kunture_%C3%89thiopie_fond.jpg) "หินคม" เป็นเครื่องมือหิน Oldowan ซึ่งพบครั้งแรกที่โกรกธาร Olduvai gorge เป็นเครื่องมือหินขั้นพื้นฐานที่สุดของมนุษย์ (ยุคหินเก่าต้น) เริ่มตั้งแต่ H. habilis[D]
"หินคม" เป็นเครื่องมือหิน Oldowan ซึ่งพบครั้งแรกที่โกรกธาร Olduvai gorge เป็นเครื่องมือหินขั้นพื้นฐานที่สุดของมนุษย์ (ยุคหินเก่าต้น) เริ่มตั้งแต่ H. habilis[D] -
 ขวานมืออะชูเลียน (Acheulean) จากเทศมณฑลเคนต์ สหราชอาณาจักร เป็นเครื่องมือหินเหล็กไฟของ H. erectus/ergaster (ยุคหินเก่าต้น)
ขวานมืออะชูเลียน (Acheulean) จากเทศมณฑลเคนต์ สหราชอาณาจักร เป็นเครื่องมือหินเหล็กไฟของ H. erectus/ergaster (ยุคหินเก่าต้น) -
 มีดหินเหล็กไฟจากวัฒนธรรม Aurignacian ประเทศฝรั่งเศส (ยุคหินเก่าปลาย)
มีดหินเหล็กไฟจากวัฒนธรรม Aurignacian ประเทศฝรั่งเศส (ยุคหินเก่าปลาย) -
 Venus of Willendorf พบในประเทศออสเตรีย เป็นตัวอย่างของศิลปะยุคหินเก่าปลาย
Venus of Willendorf พบในประเทศออสเตรีย เป็นตัวอย่างของศิลปะยุคหินเก่าปลาย
!["หินคม" เป็นเครื่องมือหิน Oldowan ซึ่งพบครั้งแรกที่โกรกธาร Olduvai gorge เป็นเครื่องมือหินขั้นพื้นฐานที่สุดของมนุษย์ (ยุคหินเก่าต้น) เริ่มตั้งแต่ H. habilis[D]](/wiki/%E0%B9%84%E0%B8%9F%E0%B8%A5%E0%B9%8C:Pierre_taill%C3%A9e_Melka_Kunture_%C3%89thiopie_fond.jpg)



เทคโนโลยี Oldowan - H. habilis/อื่น ๆ - ยุคหินเก่าต้น
แก้การใช้เครื่องมือหินมีหลักฐานชัดเจนเป็นครั้งแรกเมื่อ 2.6 ล้านปีก่อน[D] เมื่อ H. habilis ในแอฟริกาตะวันออกใช้เครื่องมือหิน Oldowan ประเภท "มีดสับเนื้อ" ที่ทำจากหินก้อนกลม ๆ (ดูรูป) ที่ทำให้แตกโดยการตีแบบง่าย ๆ[52] ซึ่งเป็นเครื่องหมายบอกการเริ่มต้นของยุคหินเก่า ซึ่งดำเนินไปจนสุดยุคน้ำแข็งสุดท้ายที่ประมาณ 10,000 ปีก่อน ยุคหินเก่าที่เริ่มที่ 2.6 ล้านปีก่อน โดยแบ่งออกเป็น 3 ช่วงคือ
- ยุคหินเก่าต้น (Lower Paleolithic) หรือยุคหินเก่าล่าง[AC]ซึ่งยุติที่ 350,000-300,000 ปีก่อน
- ยุคหินเก่ากลาง (Middle Paleolithic) ซึ่งยุติที่ 50,000-30,000 ปีก่อน
- ยุคหินเก่าปลาย (Upper Paleolithic) หรือยุคหินเก่าบน ซึ่งยุติที่ 10,000 ปีก่อน
เทคโนโลยี Acheulean - H. ergaster/erectus - ยุคหินเก่าต้น
แก้ส่วนช่วงระหว่าง 1.76 ล้าน-250,000 ปีก่อนยังรู้จักว่า สมัยเทคโนโลยี Acheulean อีกด้วย[203] เพราะเป็นช่วงที่ H. ergaster/erectus เริ่มในแอฟริกา[14] ได้ทำขวานมือมีสองคมจากจากหินเหล็กไฟและหินควอร์ตไซต์ก้อนใหญ่ (ดูรูป) โดยตอนต้นใช้หินตีส่วนขอบให้แตกออกไปจนเกิดความคม และต่อจากนั้นก็แต่งรอบ ๆ ส่วนคมให้มีรูปตามที่ต้องการ[203][14]
เทคโนโลยี Mousterian - Neanderthal (ยุคหินเก่ากลาง), H. sapiens (ยุคหินกลางแอฟริกา)
แก้หลังจาก 300,000 ปีก่อนในช่วงยุคหินเก่ากลาง เทคนิคที่ละเอียดขึ้นที่เรียกว่า Levallois technique ก็พัฒนาขึ้น (เป็นสมัย Mousterian) ซึ่งเป็นการตีหินเป็นลำดับ เป็นเทคนิคที่ใช้สร้างหัวแหลม (points), หินขูด (scraper), หินแล่ (slicer), และเข็มเจาะ (awl)[204] และเชื่อว่ามนุษย์ Neanderthal ได้พัฒนาขึ้นด้วย เพราะพบเครื่องมือหินที่เก่าแก่ที่สุดในประเทศฝรั่งเศสที่ประมาณ 300,000 ปีก่อน[205]
เทคโนโลยีนี้ก็พบในที่อื่น ๆ อีกด้วย คือที่เลแวนต์ บางส่วนของแอฟริกา เอเชียกลาง และไซบีเรีย[204][206][207] โดยในแอฟริกา เทคโนโลยีกลุ่มนี้จะเรียกว่าเป็นเครื่องมือช่วงยุคหินกลาง (Middle Stone Age) และสัมพันธ์กับมนุษย์กลุ่มอื่น ๆ ที่ไม่ใช่นีแอนเดอร์ทาล (เพราะไม่มีมนุษย์กลุ่มนี้ในแอฟริกา) เทียบกับในยุโรปที่เทคโนโลยีสัมพันธ์กับมนุษย์กลุ่มนีแอนเดอร์ทาล และเรียกว่าเป็นเครื่องมือยุคหินเก่ากลาง (Middle Paleolithic)[204] ในแอฟริกา เครื่องมือเก่าแก่สุดมีอายุ 276,000 ปีก่อน โดยการหาอายุที่น่าเชื่อถือมากที่สุด (ณ โบราณสถาน Gademotta ประเทศเอธิโอเปีย)[208]
วัฒนธรรม Aurignacian - H. sapiens - ยุคหินเก่ากลางและปลาย
แก้ประมาณระหว่าง 47,000-28,000 ปี[38] ในยุคหินเก่าปลาย ก็เกิดวัฒนธรรม Aurignacian ในยุโรปและเอเชียตะวันตกเฉียงใต้ ที่เริ่มมีการสร้างมีดหินยาว ทำเป็นมีด ใบมีด และสิ่ว[209] เป็นช่วงเวลานี้ด้วยที่เริ่มปรากฏสิ่งต่าง ๆ ดังต่อไปนี้ คือ
- ภาพศิลป์ในถ้ำ ที่เก่าที่สุดพบในภาคเหนือของประเทศสเปนมีอายุกว่า 40,800 ปี ซึ่งอาจวาดโดย H. sapiens[210] แต่นักวิทยาศาสตร์บางพวกเสนอว่าอาจวาดโดย Neanderthal[211]
- ศิลปะรูปลักษณ์ทั้งของสัตว์และมนุษย์
- ที่เก่าที่สุดของมนุษย์พบที่ประเทศเยอรมนี เป็นรูป 3 มิติชื่อว่า "Venus of Willendorf" สูง 6 ซม มีอายุ 35,000-40,000 ปีก่อน เป็นศิลปะรูปลักษณ์ของมนุษย์ที่เก่าที่สุดของโลก
- ที่เก่าของสัตว์พบที่ประเทศเยอรมนี เป็นรูป 3 มิติทำด้วยงาชื่อว่า "Lion man of the Hohlenstein Stadel" สูง 29.6 ซม มีอายุ 40,000 ปีเป็นศิลปะรูปลักษณ์ที่เก่าที่สุดของโลก[212]
- เครื่องดนตรี ที่เก่าที่สุดในโลกเป็นขลุ่ยทำด้วยงามีอายุ 42,000 ปี พบในประเทศเยอรมนี[213]
สิ่งประดิษฐ์เหล่านี้ แสดงถึงพฤติกรรมปัจจุบันของมนุษย์ที่ปรากฏชัดเจนในช่วงนี้ (ดูเพิ่มที่หัวข้อ [[#Behavioral modernity[|การเปลี่ยนมามีพฤติกรรมปัจจุบัน]]) ซึ่งเป็นเหตุให้นักวิชาการสัมพันธ์วัฒนธรรม Aurignacian กับ H. sapiens คือมนุษย์ปัจจุบัน (กลุ่มมนุษย์ต้นตระกูลคนยุโรปที่เรียกว่า Cro-Magnon) ที่สันนิษฐานว่า อพยพเข้าไปในยุโรปในช่วงเวลานี้[214][38][76][28] และทำให้เกิดการสันนิษฐานที่ยังไม่มีข้อยุติว่า มนุษย์ Neanderthal อาจจะไม่สามารถแข่งขันกับมนุษย์ปัจจุบันผู้มีเทคโนโลยีและพฤติกรรมที่ก้าวหน้ากว่าเช่นนี้ แล้วจึงทำให้สูญพันธุ์ไปเมื่อประมาณ 28,000 ปีก่อน[104] (ดูสันนิษฐานอื่น ๆ เกี่ยวกับการสูญพันธุ์ของ Neanderthal ที่หัวข้อ Neanderthal และ Denisovan)
อย่างไรก็ดี ก็ยังมีที่อื่น ๆ นอกยุโรปที่ปรากฏวัฒนธรรมคล้ายวัฒนธรรม Aurignacian อีกด้วย รวมทั้งเอเชียตะวันตกเฉียงใต้คือประเทศซีเรีย เขตปาเลสไตน์ กับประเทศอิสราเอล และเขตไซบีเรีย[207] เครื่องมือหินที่ทำเป็นมีดยาวในแอฟริกาที่เก่าที่สุดพบที่โบราณสถาน Kapthurin เคนยา และเครื่องมือแบบผสมจาก Twin Rivers แซมเบีย ทั้งสองมีอายุอาจมากถึง 300,000 ปี[38] นอกจากนั้น เริ่มตั้งแต่ปี ค.ศ. 1997 ก็เริ่มมีหลักฐานจากถ้ำ Blamboo Cave[215] และตั้งแต่ปี ค.ศ. 2000 จากแหลม Pinnacle Point[216] (ทั้งสองในประเทศแอฟริกาใต้) เป็นสิ่งประดิษฐ์ต่าง ๆ ที่นักวิชาการได้ใช้เป็นหลักฐานของ "พฤติกรรมปัจจุบัน" ของมนุษย์ แต่มีกำหนดอายุในช่วงยุคหินเก่ากลาง (100,000-70,000 ปีก่อน) ซึ่งเก่ากว่าที่พบในยุโรป ทั้งหมดเป็นหลักฐานคัดค้านทฤษฎี "Great Leap Forward" (การกระโดดก้าวไปข้างหน้า) ของพฤติกรรมปัจจุบันที่เกิดขึ้นโดยฉับพลัน[217] ที่จะกล่าวถึงต่อไป
มนุษย์กับไฟ
แก้เดิมมนุษย์มีเพียงอาหารเท่านั้นเป็นแหล่งพลังงาน ต่อมาเริ่มรู้จักใช้ไฟ ใช้แรงงานสัตว์ การเพาะปลูก จึงเริ่มมีการใช้พลังงานเพิ่มมากขึ้น แต่ยังไม่มากนัก มนุษย์เริ่มใช้พลังงานอย่างรวดเร็วในยุคปฏิวัติอุตสาหกรรม มีการประดิษฐ์เครื่องจักรกล เริ่มใช้ถ่านหิน พลังน้ำ
วิวัฒนาการการใช้พลังงาน
แก้- ยุคแรกเริ่ม(ยุคหินเก่า)มนุษย์ดำรงชีวิตแบบง่าย ๆ อยู่ตามธรรมชาติ
- ยุคล่าสัตว์(ยุคหินใหม่)มีการรวมกลุ่มเพื่อล่าสัตว์ รู้จักใช้ไฟล่าสัตว์
- ยุคอุตสาหกรรมก้าวหน้า หลังสงครามโลกครั้งที่ 1 มนุษย์ใช้เชื้อเพลิงน้ำมัน ถ่านหินและใช้เทคโนโลยีมากขึ้นตสาหกรรมเริ่มต้น มีการประดิษฐ์เครื่องจักรไอน้ำ มนุษย์เริ่มรู้จักใช้เชื้อเพลิง
มนุษย์ รู้จักใช้ไฟเมื่อมีการระเบิดของภูเขาไฟในอัฟริกา ซึ่งเกิดขึ้นเมื่อประมาณ 4 แสนปีมาแล้ว นับเป็นการค้นพบทางวิทยาศาสตร์ครั้งแรกของมนุษย์ หลังจากนั้นความเป็นอยู่ของมนุษย์ก็เริ่มดีขึ้น และไฟถูกนำมาใช้ประโยชน์สืบต่อกันมาจนทุกวันนี้[218] มนุษย์จุดไฟได้ครั้งแรกเมื่อ 4 แสนปีก่อน และช่วยให้มนุษย์อพยพจากแอฟริกามุ่งหน้าสู่ยุโรปได้ มนุษย์ในยุคแรกๆ ซึ่งอาจจะเป็นในช่วงของ โฮโม อิเร็คตัส (Homo erectus) หรือโฮโม เออร์กัสเตอร์ (Homo ergaster) นั้น ได้จัดการและใช้ประโยชน์จากไฟ แต่ก็ยังไม่ชัดเจนว่า มนุษย์โบราณสามารถจุดไฟได้เองหรือขโมยมาจากการเกิดขึ้นเองตามธรรมชาติ อย่างปรากฏการณ์ฟ้าผ่า จากการวิเคราะห์หินไฟ (flint) บริเวณพื้นที่ทางโบราณคดีตรงชายฝั่งแม่น้ำจอร์แดน รอยเตอร์ระบุว่าทีมนักวิจัยจากมหาวิทยาลัยฮิบรู (Hebrew University) เยรูซาเลม อิสราเอล ได้พบอารยธรรมเริ่มต้น ซึ่งมีการเรียนรู้ที่จะจุดไฟ และนั่นกลายเป็นจุดเปลี่ยนที่ทำให้มนุษย์โบราณได้ท่องผจญภัยในดินแดนที่ไม่รู้จัก[219]
การเปลี่ยนมามีพฤติกรรมปัจจุบัน
แก้จนกระทั่งถึงประมาณ 50,000-40,000 ปีก่อน การใช้เครื่องมือหินดูเหมือนจะมีการพัฒนาเป็นขั้น ๆ โดยในแต่ละก้าว H. habilis, H. ergaster/erectus, และ H. neanderthalensis จะเริ่มต้นตรงที่สูงขึ้นจากขั้นก่อน ๆ แต่เมื่อเริ่มแล้ว การพัฒนาต่อ ๆ ไปจะเป็นไปอย่างล่าช้า ในปัจจุบัน นักบรรพมานุษยวิทยายังพิจารณากันอยู่ว่า สปีชีส์ของ Homo เหล่านี้ มีลักษณะทางวัฒนธรรมหรือพฤติกรรมบางอย่างหรือหลายอย่าง ที่เหมือนกับมนุษย์ปัจจุบันเช่น ภาษา ความคิดทางสัญลักษณ์ที่ซับซ้อน ความคิดสร้างสรรค์ทางเทคโนโลยี ฯลฯ หรือไม่ เพราะว่า มนุษย์เหล่านั้นดูเหมือนว่า จะค่อนข้างอนุรักษนิยมในเรื่องวัฒนธรรม เพราะรักษาไว้ซึ่งการใช้เทคโนโลยีแบบง่าย ๆ และรูปแบบการแสวงหาอาหารเป็นช่วงระยะเวลายาวนาน
ประมาณ 50,000 ปีก่อน วัฒนธรรมของมนุษย์ปัจจุบันจึงปรากฏหลักฐานว่าเริ่มพัฒนาเร็วขึ้น การเปลี่ยนแปลงมามีพฤติกรรมปัจจุบันได้รับการกำหนดว่าเป็น "Great Leap Forward" (การกระโดดก้าวไกลไปข้างหน้า) ของคนยูเรเชีย[220] หรือเป็น "Upper Palaeolithic Revolution" (ปฏิวัติยุคหินเก่าปลาย)[221] เพราะการปรากฏอย่างฉับพลันของหลักฐานทางโบราณคดี ที่บ่งถึงพฤติกรรมปัจจุบันของมนุษย์ แต่ว่า นักวิชาการบางท่านก็พิจารณาการเปลี่ยนแปลงเช่นนี้ว่า เป็นไปอย่างค่อยเป็นค่อยไป โดยได้ปรากฏขึ้นแล้วบ้างในสายมนุษย์โบราณแอฟริกาช่วง 200,000 ปีก่อน[222][223] ดังที่แสดงโดยหลักฐานจากถ้ำ Blamboo Cave ในประเทศแอฟริกาใต้[215]
ในช่วงนี้ มนุษย์ปัจจุบันเริ่มฝังคนตาย[AB] ใช้เสื้อผ้า[AA] ล่าสัตว์ด้วยเทคนิคที่ซับซ้อนขึ้น (เช่นใช้หลุมพรางเป็นต้น)[14] และทำงานศิลป์เกี่ยวกับภาพวาดในถ้ำ[210]
เมื่อวัฒนธรรมเจริญขึ้น มนุษย์กลุ่มต่าง ๆ ก็เริ่มสร้างสิ่งใหม่ ๆ เสริมเข้ากับเทคโนโลยีที่มีอยู่แล้ว สิ่งประดิษฐ์เช่น เบ็ดปลา ลูกกระดุม และเข็มกระดูก มีความแตกต่างกันระหว่างมนุษย์กลุ่มต่าง ๆ ซึ่งเป็นอะไรที่ไม่เคยเห็นในวัฒนธรรมมนุษย์ก่อน 50,000 ปีก่อน คือโดยทั่ว ๆ ไปแล้ว กลุ่มต่าง ๆ ของ H. neanderthalensis ไม่ค่อยมีความแตกต่างกันทางเทคโนโลยี
ในบรรดาตัวอย่างที่สามารถหาหลักฐานทางโบราณคดีได้ของพฤติกรรมมนุษย์ปัจจุบัน นักมานุษยวิทยารวมเอา[38][232]
- การทำเครื่องมือเฉพาะกิจ เครื่องมือผสม หรือเครื่องมือที่ทำได้อย่างละเอียดละออ (เช่นเครื่องมือหินที่ประกอบกับด้าม)
- อาวุธที่ใช้ยิงได้ (เช่นธนู)
- การเล่นเกมและดนตรี
- เรือหรือแพ
- เครื่องใช้ดินเผา
- การก่อสร้างและการจัดระเบียบของที่อยู่
- เทคนิคการล่าสัตว์ที่ซับซ้อน
- การตกปลา และการจับสัตว์เล็ก ๆ เช่นกระต่ายเป็นต้น
- การหุงอาหารและการปรุงอาหาร คือไม่ได้ทานอย่างดิบ ๆ
- การเก็บรักษาอาหารเป็นเสบียง
- สัตว์เลี้ยง
- เครือข่ายการแลกเปลี่ยนสินค้า
- การไปได้ในที่ไกล ๆ
- ศิลปะรูปลักษณ์คือรูปสัตว์และรูปมนุษย์เป็นต้น (เช่นภาพวาดในถ้ำ)
- การใช้สี เช่นสีดิน และเครื่องประดับเพื่อการประดับตกแต่งที่อยู่หรือร่างกาย
- การฝังศพพร้อมกับวัตถุต่าง ๆ
แต่ว่า การหาข้อยุติก็ยังเป็นไปว่า มี "การปฏิวัติ" ที่เกิดโดยฉับพลันที่นำมาสู่พฤติกรรมของมนุษย์ปัจจุบัน หรือว่า เป็นพัฒนาการที่เกิดขึ้นอย่างค่อยเป็นค่อยไปเริ่มต้นตั้งแต่ 250,000-300,000 ปีก่อน[38] นอกจากนั้นแล้ว ก็ยังมีนักวิชาการที่คัดค้านด้วยว่า ตัวอย่างพฤติกรรมปัจจุบันให้ความสำคัญกับหลักฐานที่มีในแอฟริกาน้อยเกินไป (ซึ่งหายากกว่าเก็บรักษาไว้ได้ในสภาพที่แย่กว่าเพราะภูมิอากาศ) หรือคลุมเครือเกินไปเพราะว่าสามารถอธิบายได้โดยเหตุผลอื่นที่ง่ายกว่า หรือมีปัญหาในการใช้วิธีและมาตรฐานทดสอบเดียวกันในช่วงสมัยต่าง ๆ กัน[233] นอกจากนั้นแล้ว ก็เริ่มจะปรากฏหลักฐานที่คัดค้านทฤษฎี "Great Leap Forward" (การกระโดดก้าวไปข้างหน้า) คือพบสิ่งประดิษฐ์ที่ใช้เป็นหลักฐานแสดงพฤติกรรมปัจจุบันของมนุษย์ในแอฟริกาใต้ในช่วงยุคหินเก่ากลาง ที่เก่าแก่กว่าที่พบยุโรป[216]
ส่วนคำว่า Behavioral modernity (แปลว่า การมีพฤติกรรมปัจจุบัน) เป็นบัญญัติภาษาอังกฤษที่ใช้ในมานุษยวิทยา โบราณคดี และสังคมวิทยาเมื่อกล่าวถึงลักษณะสืบสายพันธุ์ (trait) ที่ทำมนุษย์ปัจจุบันและบรรพบุรุษเมื่อไม่นานนี้ให้ต่างจากไพรเมตที่ยังมีชีวิตอื่น ๆ และจากสายพันธุ์มนุษย์ที่สูญพันธุ์ไปแล้ว เป็นจุดเริ่มต้นที่มนุษย์สปีชีส์ Homo sapiens เริ่มปรากฏความสามารถในการใช้ความคิดเชิงนามธรรม (ความคิดเชิงสัญลักษณ์ คือใช้ตัวแทนที่หมายถึงสิ่งอื่น) และเริ่มแสดงความคิดสร้างสรรค์ทางด้านวัฒนธรรม พัฒนาการเหล่านี้เชื่อว่ามีความสัมพันธ์เกี่ยวกับกำเนิดภาษา[234] องค์ประกอบของพฤติกรรมปัจจุบันอาจจะรวมถึง เครื่องมือที่ทำได้อย่างละเอียด การตกปลา การแลกเปลี่ยนสินค้าระหว่างกลุ่มที่ไกลกัน การประดับตัว ศิลปะเชิงรูปลักษณ์ เกม (การละเล่น) ดนตรี การหุงอาหาร และพิธีการฝังศพ
วิวัฒนาการของมนุษย์ในเร็ว ๆ นี้และในปัจจุบัน
แก้การคัดเลือกโดยธรรมชาติก็ยังเป็นไปในมนุษย์ปัจจุบันกลุ่มต่าง ๆ ยกตัวอย่างเช่น ในมนุษย์กลุ่มที่มีความเสี่ยงสูงต่อโรคที่ทำให้อ่อนกำลังอย่างรุนแรงคือโรคคูรุ[AD] ปรากฏคนที่มียีนพรีออน "G127 V" ซึ่งเป็นอัลลีลเป็นยีนรูปแปรที่มีภูมิคุ้มกันต่อโรคมากกว่าอัลลีลที่ไม่มีภูมิคุ้มกัน ความชุกของการกลายพันธุ์ (หรือรูปแปรทางกรรมพันธุ์) เช่นนี้ เกิดขึ้นเพราะการรอดชีวิตของบุคคลผู้มีภูมิต้านทานโรค[236][237] วิวัฒนาการที่พบในมนุษย์กลุ่มอื่น ๆ รวมทั้งวัยเจริญพันธุ์ที่มีระยะยาวขึ้น บวกกับการลดระดับของคอเลสเตอรอล กลูโคสในเลือด และความดันเลือด[238]
มีการเสนอว่า วิวัฒนาการของมนุษย์ได้เร่งเร็วขึ้นตั้งแต่เกิดการพัฒนาทางเกษตรกรรมและอารยธรรมเมื่อ 10,000 ปีก่อน และนี่ทำให้เกิดความแตกต่างทางกรรมพันธุ์อย่างสำคัญในกลุ่มต่าง ๆ ของมนุษย์[239] การมีเอนไซม์แล็กเทสในวัยผู้ใหญ่ (Lactase persistence) คือการที่ยังมีการทำงานของเอนไซม์แล็กเทสเพื่อย่อยแล็กโทสซึ่งเป็นน้ำตาลในนมตลอดจนถึงวัยผู้ใหญ่ เป็นอีกตัวอย่างหนึ่งของวิวัฒนาการที่เกิดขึ้นเร็ว ๆ นี้ เพราะว่าโดยปกติของสปีชีส์ต่าง ๆ ในสัตว์เลี้ยงลูกด้วยนม การทำงานของเอนไซม์จะลดลงอย่างสำคัญหลังจากเลิกนม[240] ประชากรในประเทศต่าง ๆ มียีนผลิตเอนไซม์แล็กเทสในวัยผู้ใหญ่เป็นเปอร์เซ็นต์ที่ไม่เท่ากัน คือเริ่มตั้งแต่ชาวดัตช์ที่ 99%[241] จนถึงชาวไทยที่ 2%[242] และกลุ่มชนพื้นเมืองอเมริกันในสหรัฐอเมริกาที่ 0%[242] ซึ่งส่วนสัดเช่นนี้ขึ้นอยู่กับว่าเป็นกลุ่มประชากรที่บริโภคนมในวัยผู้ใหญ่เป็นเวลามายาวนานในประวัติหรือไม่
อย่างไรก็ดี วิวัฒนาการของมนุษย์ที่ปรากฏเร็ว ๆ นี้ดูเหมือนจะจำกัดอยู่ในเรื่องภูมิต้านทานต่อโรคติดต่อโดยมาก เป็นโรคที่มนุษย์ติดข้ามสปีชีส์มาจากสัตว์เลี้ยง[39] แต่เพราะไร้เหตุกดดันทางการคัดเลือกโดยธรรมชาติ หรือเพราะไร้โอกาสที่จะเกิดสปีชีส์ใหม่ เช่นโอกาสการอยู่แยกต่างหากโดยไม่ติดต่อคนอื่นนอกพื้นที่ วิวัฒนาการของมนุษย์เร็ว ๆ นี้ (รวมที่ปรากฏและไม่ปรากฏ) โดยมากก็จะเป็นการเปลี่ยนความถี่ยีนอย่างไม่เจาะจง (genetic drift) นอกจากนั้นแล้ว ยังปรากฏอีกด้วยว่า ทั้งมนุษย์ทั้งวงศ์ลิงใหญ่แอฟริกัน (รวมกอริลลาและชิมแปนซี) ปรากฏการวิวัฒนาการที่ช้าลงจากลิงสายพันธุ์อื่น ๆ ซึ่งอาจเกิดขึ้นเพราะแต่ละชั่วอายุมีความยาวนานยิ่งขึ้น[10]
ประมวลข้อมูลมนุษย์สปีชีส์ต่าง ๆ
แก้- ลำดับการเกิดขึ้นของสายพันธุ์มนุษย์ ดู แผนภูมิสปีชีส์ตามกาลเวลา
- กาลานุกรมวิวัฒนาการมนุษย์เริ่มตั้งแต่ 4,000 ล้านปีก่อน
- ตารางเปรียบเทียบสปีชีส์ต่าง ๆ ของมนุษย์สกุลโฮโม
- รายการซากดึกดำบรรพ์สายพันธุ์มนุษย์
แผนภาพแสดงความใกล้เคียงกันของสายพันธุ์มนุษย์
แก้แผนภาพวิวัฒนาการแบบแคลดิสติกส์ของสายพันธุ์มนุษย์ (คือแผนภาพแสดงความใกล้เคียงกันของสายพันธุ์มนุษย์ อังกฤษ: hominin cladogram) เป็นแผนภาพแสดงการจัดกลุ่มของสัตว์ในสายพันธุ์มนุษย์ (ที่แยกออกจากลิงใหญ่แอฟริกันแล้ว) โดยลักษณะเฉพาะที่มีร่วมกันอย่างหนึ่งหรือมากกว่าหนึ่งของกลุ่มนั้น ๆ ที่สืบมาจากบรรพบุรุษที่มีร่วมกัน และไม่พบในกลุ่มอื่น ๆ ดังนั้น จึงพิจารณาว่ามนุษย์ที่อยู่ในกลุ่มเดียวกันจะมีประวัติการสืบสายพันธุ์ร่วมกันและมีความใกล้ชิดต่อกันและกัน มากกว่าต่อสมาชิกในกลุ่มอื่น ๆ[243][244]
ในบทความนี้แสดงสามแบบ ให้สังเกตว่า สปีชีส์ที่ "อยู่ในกลุ่มเดียวกัน" ก็คือสปีชีส์ที่มีจุดเริ่มร่วมกันนั่นเอง
| ตาม Wood & Richmond (ค.ศ. 2000) เป็นการวิเคราะห์ใช้แต่ข้อมูลหัวกะโหลกและฟัน[11] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| ตาม Strait & Grine (ค.ศ. 2004) เป็นการวิเคราะห์ใช้แต่ข้อมูลหัวกะโหลกและฟัน[245] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| ตาม Organ et al. (ค.ศ. 2011)[246] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
รายชื่อสปีชีส์
แก้รายชื่อนี้เป็นไปตามลำดับเวลาจากซ้ายไปขวาโดยสกุล ให้แทนอักษรย่อในทวินามด้วยชื่อสกุล
|
|
|
ดูเพิ่ม
แก้เชิงอรรถ
แก้- ↑ 1.0 1.1 1.2 1.3 1.4 ให้สังเกตว่า โดยมาก ช่วงเวลาที่ให้เกี่ยวกับการแยกสายพันธุ์มักจะมาจากข้อมูลทางดีเอ็นเอ แต่ว่า ข้อมูลจากงานต่าง ๆ มักจะให้เวลาที่ไม่เหมือนกันขึ้นอยู่กับตัวแปรต่าง ๆ ของงานเช่นแบบจำลองของวิวัฒนาการที่ใช้เป็นต้น ดังนั้น ข้อมูลทางดีเอ็นเอนี้ควรจะเป็นช่วงตัวเลขเพื่อให้ตรงกับผลการทดลองโดยทั่ว ๆ ไป แต่ในบางที่ที่ไม่สำคัญ การใช้ตัวเลขเดี่ยว ๆ นั้นเป็นความสะดวก ดูตัวอย่างความหลากหลายของเวลาการแยกสายพันธุ์ของ Hylobatidae จาก Hominidae ที่ Time Tree[6]
- ↑ 2.0 2.1 2.2 เผ่า Hominini (อังกฤษ: hominin) ปกติใช้เรียกสายพันธุ์ของมนุษย์ที่ลิงชิมแปนซีได้แยกออกไปแล้ว ซึ่งในที่สุดกลายเป็นมนุษย์สกุล Homo
- ↑ 3.0 3.1 3.2 มีหลักฐานใหม่ที่แสดงว่า H. habilis อาจเริ่มวิวัฒนาการขึ้นตั้งแต่ 2.8 ล้านปีก่อน ดูรายละเอียด
- ↑ 4.0 4.1 4.2 4.3 4.4 4.5 4.6 4.7 หลักฐานการใช้เครื่องมือหินที่เก่าที่สุดมีอายุประมาณ 3.4 ล้านปีก่อน (สมัยไพลโอซีนปลาย) คือ มีการพบกระดูกที่มีรอยตัดและรอยทุบในประเทศเอธิโอเปียใกล้กับซากของทารก homonin สปีชีส์ Australopithecus afarensis เหมือนกับลูซี่ แต่ว่าเครื่องมือหินเทคโนโลยีแบบ Oldowan ที่เก่าที่สุดมีอายุ 2.6 ล้านปีก่อน ในสมัยไพลโอซีนปลาย (ยุคหินเก่าต้น) พบที่เอธิโอเปีย ซึ่งเป็นช่วงก่อนการเกิดขึ้นของ H. habilis ซึ่งเป็นสกุลที่นักโบราณคดีสัมพันธ์กับการใช้เทคโนโลยีนี้มากที่สุด หลังจากนี้ เทคโนโลยี Oldowan ก็ได้กระจายไปทั่วแอฟริกา แต่ว่านักโบราณคดียังไม่แน่ใจว่า hominin สกุลไหนสร้างเทคโนโลยีนี้ขึ้นมาก่อน บางพวกเชื่อว่าเป็น Australopithecus garhi บางพวกเชื่อว่าเป็น Homo habilis[14] แต่ H. habilis เป็นพวกที่ใช้เทคโนโลยีนี้มากที่สุดในแอฟริกา จนกระทั่งถึง 1.9 ถึง 1.8 ล้านปีก่อน ที่ H. ergaster/erectus สืบทอดเทคโนโลยีนี้ต่อมา แม้ว่า เทคโนโลยีนี้จะพบในแอฟริกาใต้และตะวันออกในช่วง 2.6 ถึง 1.7 ล้านปีก่อน แต่ก็มีการกระจายออกไปในยูเรเชียพร้อมกับการอพยพออกจากแอฟริกาของ H. ergaster/erectus ซึ่งนำเทคโนโลยีนี้ไปทางทิศตะวันออกไปจนถึงเกาะชวา (อินโดนีเซีย) เมื่อประมาณ 1.8 ล้านปีก่อน และไปถึงจีนภาคเหนือเมื่อ 1.6 ล้านปีก่อน
- ↑ 5.0 5.1 5.2 5.3 5.4 5.5 การขยายขนาดของสมอง (อังกฤษ: encephalization) เป็นการขยายขนาดของสมองเมื่อเทียบกับตัว คือสัตว์ที่มีร่างกายใหญ่โตต้องมีสมองที่ใหญ่กว่า ดังนั้น การขยายขนาดสมองของมนุษย์จริง ๆ แล้วเป็นการขยายขนาดของสมองในระดับที่สูงกว่าการขยายขนาดของตัว
- ↑ ในมนุษย์ SRGAP2 เป็นยีนที่เข้ารหัสโปรตีน "SLIT-ROBO Rho GTPase-activating protein 2"[18][19]
เป็นหนึ่งใน 23 ยีนที่เกิดการก๊อปปี้ในมนุษย์ที่ไม่ปรากฏในไพรเมตประเภทอื่น[20]
คือเกิดการทำซ้ำถึงสามครั้งในจีโนมมนุษย์ภายใน 3.4 ล้านปีที่ผ่านมา ชุดแรกเรียกว่า SRGAP2B ที่ 3.4 ล้านปีก่อน ชุดที่สองเรียกว่า SRGAP2C ที่ 2.4 ล้านปีก่อน และชุดสุดท้ายเรียกว่า SRGAP2D ที่ 1 ล้านปีก่อน
ส่วนยีน SRGAP2 เป็นชุดที่มีในสัตว์เลี้ยงลูกด้วยนมทั้งหมด (เป็นชุดที่ในมนุษย์เรียกว่า SRGAP2A) และชุด SRGAP2C ทำให้เกิดการแสดงออกเป็นโปรตีนแบบสั้นที่มีในมนุษย์ปัจจุบันทั้งหมด[21]
เป็นโปรตีนที่มีหน้าที่ยับยั้งการแสดงออกของ SRGAP2A ที่เกิดขึ้นก่อน ทำให้เกิดผลสองอย่างคือ[22]
- ช่วยให้นิวรอนเคลื่อนไปจากแหล่งกำเนิดสู่จุดเป้าหมายได้เร็วขึ้น
- ยืดระยะเวลาที่ไซแนปส์จะถึงจุดสูงสุดในการพัฒนา และเพิ่มความหนาแน่นของไซแนปส์ในเปลือกสมอง (cerebral cortex)
- ↑ 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 7.8 ในงานปี 2560 การหาอายุของเครื่องมือหินในชั้นตะกอนเดียวกับซากดึกดำบรรพ์มนุษย์ Homo sapiens ที่เก่าแก่ที่สุด ณ โบราณสถาน Jebel Irhoud ประเทศโมร็อกโก ได้ใช้เทคนิค thermoluminescence
ซึ่งสามารถหาเวลาที่ผ่านไปหลังจากเผาแร่ผลึกในเครื่องมือหินเป็นครั้งสุดท้าย โดยได้อายุต่าง ๆ 14 รายการ[197]
และมีค่าเฉลี่ยถ่วงน้ำหนักที่ 315,000 ± 34,000 ปี[191]
เทียบกับอายุราว 195,000 ปีของซาก Omo remains โดยเพิ่มอายุเก่าแก่ที่สุดของซากมนุษย์ที่เคยขุดพบราว 100,000 ปี จาก 200,000 ปี เป็น 300,000 ปี[193][194][195][196][198]
- ดูข้อมูลเพิ่มเติมที่หัวข้อซากมนุษย์ H. sapiens/"ที่มีกายวิภาคปัจจุบัน" ที่เก่าแก่ที่สุด
- ↑ 8.0 8.1 8.2 8.3 Genetic admixture เกิดขึ้นเมื่อสัตว์หรือบุคคลจากกลุ่มประชากรที่ก่อนหน้านี้อยู่แยกขาดจากกันเกิดการผสมพันธุ์กัน มีผลเป็นการสร้างสายพันธุ์ใหม่ในประชากร ซึ่งบางครั้งมีผลเป็นการทำการปรับตัวต่อสิ่งแวดล้อมในพื้นที่ (local adaptation) ให้เนิ่นช้า เพราะว่าได้รับเอาลักษณะทางพันธุกรรม (genotype) ที่ไม่ได้มีการปรับตัว ที่ไม่เหมือนกับของบุคคลในพื้นที่ มีผลเป็นการป้องกันการเกิดสปีชีส์ใหม่เพราะทำกลุ่มต่าง ๆ ให้เหมือนกัน
- ↑ Symbolic culture (แปลว่า วัฒนธรรมสัญลักษณ์) เป็นบัญญัติที่ใช้โดยนักโบราณคดี[33] นักมานุษยวิทยาเชิงสังคม[34] และนักสังคมวิทยา[35] เพื่อทำวัฒนธรรมของมนุษย์สปีชีส์ Homo sapiens ให้แตกต่างจากวัฒนธรรมทั่ว ๆ ไปที่สัตว์อื่น ๆ มากมายมี การมีวัฒนธรรมสัญลักษณ์ไม่ใช่เป็นเพียงแต่ความสามารถในการสืบทอดวัฒนธรรมประเพณีจากรุ่นหนึ่งไปสู่อีกรุ่นหนึ่งเท่านั้น แต่ต้องมีการบัญญัติสิ่งที่จริง ๆ แล้วไม่มีในโลก แต่ปรากฏเหมือนเป็นสิ่งที่มีจริง ๆ ในวัฒนธรรมนั้น ๆ ยกตัวอย่างคือ ความดีความชั่ว พระเป็นเจ้าและนรก และบัญญัติที่เป็นที่ยอมรับของสังคมอื่น ๆ เช่นเงินตราและระบบประชาธิปไตย[36]
- ↑ มีที่อ้างอิงที่แสดงว่า Homininae แบ่งออกเป็น 3 เผ่า คือ Gorillini, Panini, และ Hominini (ดังนั้น สกุล Pan จึงอยู่ใต้เผ่า Panini และ Homo จึงอยู่ใต้เผ่า Hominini)[11][43]
- ↑ ตัวอย่างเช่น วิวัฒนาการของมนุษย์เป็นความจริงที่ได้รับการพิสูจน์มาจากวิทยาศาสตร์สาขาต่าง ๆ อย่างชัดเจน ดูหลักฐานจากศาสตร์ต่าง ๆ ที่เว็บไซต์ (ภาษาอังกฤษ) นี้[62]
- ↑ 12.0 12.1 บรรพมานุษยวิทยา (อังกฤษ: Paleoanthropology) มาจากภาษากรีกแปลว่า การศึกษามนุษย์โบราณ เป็นการรวมการศึกษาของสาขาบรรพชีวินวิทยาและมานุษยวิทยากายภาพ (physical anthropology) เข้าด้วยกัน เป็นการศึกษามนุษย์โบราณตามหลักฐานซากดึกดำบรรพ์ของสัตว์ในเผ่าย่อย homonini เช่นศึกษากระดูกที่กลายเป็นหิน หรือรอยเท้าที่สัตว์สายพันธุ์มนุษย์ได้ทิ้งไว้ เป็นการศึกษาวัฒนธรรม สังคม และชีวภาพของมนุษย์ เพื่อหาต้นกำเนิดของลักษณะทางกายภาพและพฤติกรรมที่สืบมาทางสายพันธุ์[13]
- ↑ คือ ในช่วงนั้น นักวิทยาศาสตร์เชื่อว่า ลิงกอริลลาและลิงชิมแปนซีเป็นไพรเมตที่มีความสัมพันธ์ใกล้ชิดกันมากกว่ากับมนุษย์ และประมาณว่า สายพันธุ์ของลิงเหล่านั้นแยกออกจากสายมนุษย์เมื่อประมาณ 15 ถึง 28 ล้านปีก่อน เมื่อมีการค้นพบลิงใหญ่สกุล Ramapithecus มีอายุ 14 ล้านปี ซึ่งมีลักษณะบางอย่างคล้ายมนุษย์ จึงเข้าใจว่า สกุลนี้อยู่ในสายพันธุ์ของมนุษย์ และสกุลนี้เป็นหลักฐานยืนยันเวลาของการแยกสายพันธุ์ของมนุษย์ (ปี ค.ศ. 1966) ต่อมาเมื่อมีหลักฐานทางกรรมพันธุ์เกี่ยวกับเวลาการแยกสายพันธุ์ของมนุษย์ และหลักฐานทางซากดึกดำบรรพ์อื่น ๆ เพิ่มขึ้น จึงปรากฏชัดเจนขึ้นว่า Ramapithecus เป็นบรรพบุรุษของลิงอุรังอุตัง ไม่ใช่เป็นบรรพบุรุษของมนุษย์ (ปี ค.ศ. 1982)[10]
- ↑ Molecular evolution (แปลว่า วิวัฒนาการทางโครงสร้างโมเลกุล หรืออณูวิวัฒนาการ) เป็นการเปลี่ยนแปลงลำดับของโมเลกุลในเซลล์เช่น ดีเอ็นเอ อาร์เอ็นเอ และโปรตีน ในระยะเวลายาว
ศาสตร์นี้มุ่งหมายจะอธิบายการเปลี่ยนแปลงทางชีวภาพที่ระดับโมเลกุลและระดับเซลล์
โดยใช้หลักของศาสตร์ชีววิทยาเชิงวิวัฒนาการ (evolutionary biology) และพันธุศาสตร์ประชากร (population genetics)
ประเด็นการศึกษารวมทั้ง
- อัตราการเปลี่ยนแปลง และผลการเปลี่ยนแปลง แบบนิวคลีโอไทด์เดี่ยว (single nucleotide)
- วิวัฒนาการที่ไร้ผล (neutral evolution) เทียบกับ การคัดเลือกโดยธรรมชาติ (คือวิวัฒนาการที่มีผลบวก)
- กำเนิดของยีนใหม่ ๆ
- ธรรมชาติของยีนที่ทำให้เกิดลักษณะสืบสายพันธุ์ (trait) ที่ซับซ้อน
- ธรรมชาติของยีนที่ทำให้เกิดสปีชีส์ขึ้นมาใหม่
- วิวัฒนาการของพัฒนาการ (evolution of development)
- อิทธิพลของกระบวนการวิวัฒนาการต่อความเปลี่ยนแปลงในจีโนมและลักษณะทางพันธุกรรม
- ↑ ข้อมูลวิถีการอพยพมาจากงานวิจัยเกี่ยวกับดีเอ็นเอของไมโทคอนเดรีย เส้นขาดแสดงทางอพยพที่ยังเป็นสมมติฐาน
ตัวเลขบ่งโดยสีมีหน่วยเป็นพันปีก่อน (kya)
เส้นสีน้ำเงินเป็นเขตที่คลุมไปด้วยน้ำแข็งหรือทุ่งทุนดรา (ทุ่งในอากาศหนาวเย็น มีต้นไม้บางประเภทได้เท่านั้น) ในช่วงยุคน้ำแข็งยุคก่อน
ส่วนอักษรต่าง ๆ เป็น haplogroup ของดีเอ็นเอนของไมโทคอนเดรีย (mtDNA) ซึ่งสืบสายมาจากมารดาเพียงเท่านั้น
และสามารถใช้กำหนดกลุ่มชนต่าง ๆ โดยกรรมพันธุ์ ที่มักจะเป็นไปตามเขตภูมิภาค
ยกตัวอย่างเช่น นี้เป็นการจำแนก mtDNA haplogroup โดยทั่วไป คือ
คนแอฟริกา (L, L1, L2, L3) คนตะวันออกใกล้ (J, N) คนยุโรปใต้ (J, K) คนยุโรปทั่วไป (H, V) คนยุโรปเหนือ (T, U, X) คนเอเชีย (A, B, C, D, E, F, G โดยที่ M หมายถึง C, D, E, G) และ คนพื้นเมืองอเมริกัน (A, B, C, D, และบางครั้ง X)
หลักฐานอ้างอิงเดิม (Behar et al. 2008, Gonder et al. 2007, Reed and Tishkoff) น่าจะหมายถึงงานต่อไปนี้
- Behar, Doron M.; Villems, Richard; Soodyall, Himla; Blue-Smith, Jason; และคณะ (2008). "The Dawn of Human Matrilineal Diversity". The American Journal of Human Genetics. 82 (5): 1130–1140. doi:10.1016/j.ajhg.2008.04.002. PMC 2427203. PMID 18439549. (08) 00255-3.pdf Full Article PDF (753 KB)
- Gonder, Mary Katherine; Mortensen, Holly M.; Reed, Floyd A.; de Sousa, Alexandra; และคณะ (2007). "Whole-mtDNA Genome Sequence Analysis of Ancient African Lineages". Molecular Biology and Evolution. 24 (3): 757–768. doi:10.1093/molbev/msl209. ISSN 1537-1719.
- ไม่ชัดเจนว่างานของ Reed and Tishkoff ควรจะเป็นอันไหน
- ↑ คือ เมื่อมีการอพยพย้ายถิ่นฐานออกจากที่ที่หนึ่ง ผู้ที่อพยพออกมีส่วนน้อยกว่าส่วนที่ดำรงอยู่ (founder event) ดังนั้นความหลายหลากของจีโนมจะมีอยู่ในระดับที่สูงกว่าในกลุ่มประชากรที่เป็นต้นกำเนิดของการอพยพ งานวิจัยในปี ค.ศ. 2009 พบว่า มีระดับความหลายหลากสูงในกลุ่มประชากรแอฟริกา, ยุโรปและตะวันออกกลาง, อินเดีย, เอเชียตะวันออก, โอเชียเนีย, และคนพื้นเมืองอเมริกัน ไปตามลำดับ ซึ่งโดยคร่าว ๆ แล้ว เป็นลำดับเวลาการตั้งถิ่นฐานในที่ต่าง ๆ ทั่วโลกของมนุษย์ปัจจุบัน งานทดลองนี้จึงสนับสนุนว่า แอฟริกาเป็นแหล่งกำเนิดของมนุษย์ที่อพยพไปในที่ต่าง ๆ
- ↑ นี้เป็นคำบรรยายประกอบภาพการหมุนตัวของทารกในเวลาคลอด เบื้องต้น ทารกหันหน้าเยื้องไปทางตะโพกของแม่ด้านใดด้านหนึ่ง (แถวบนสุด) เมื่อกล้ามเนื้อมดลูกของแม่ดันทารกไปถึงฐานเชิงกราน คางของทารกก็จะเริ่มไปชิดกันกับหน้าอก เมื่อศีรษะที่ชิดกับหน้าอกมาจรดฐานเชิงกรานแล้ว ก็จะเกิดการหมุนศีรษะ (internal rotation) โดยไม่ได้หมุนตัวหรือไหล่ ให้หันหน้าไปทางด้านทวารหนักของแม่ (แถวที่ 2 ซ้าย) เมื่อศีรษะโผล่ออกจากช่องคลอดแล้ว ก็จะเกิดการหมุนศีรษะกลับไปตรงกับแนวตัวและไหล่อีก (restitution) และต่อจากนั้นก็จะมีการหมุนไหล่ให้ด้านหนึ่งอยู่บนด้านหนึ่งอยู่ล่างพร้อมทั้งศีรษะซึ่งหันหน้าไปทางต้นขาของแม่ (external rotation แถว 2 ขวา) และเมื่อหมอช่วยให้ไหล่ด้านบนออกมาได้แล้ว ก็จะช่วยให้ไหล่ด้านล่างออกมาได้ต่อไป (แถว 3 ซ้าย)[92][93][94] ดูภาพยนตร์ (ไม่มีเสียงบรรยาย) จำลองการคลอดที่[95]
- ↑ 18.0 18.1 ให้สังเกตว่า สำหรับสกุล Propliopithecus และ Aegyptopithecus นักวิชาการบางพวกเชื่อว่าเป็นบรรพบุรุษร่วมกันของลิงโลกเก่าและเอป (รวมทั้งมนุษย์)[13] ส่วนบางพวกเชื่อว่า เป็นเอปยุคต้น ๆ (คือลิงโลกเก่าได้แยกสายพันธุ์ออกไปแล้ว ไม่เป็นบรรพบุรุษของลิงโลกเก่า)[115][114] ข้อมูลในที่นี้จึงเหมือนกับจะขัดแย้งกัน เพราะว่าแล้วแต่ว่าใช้หลักฐานไหน แต่ว่า นี้เป็นปัญหาโดยทั่ว ๆ ไปของการวิเคราะห์ซากดึกดำบรรพ์ที่เกิดใกล้ ๆ ช่วงระยะเวลาที่มีการแยกสายพันธุ์ คือนักวิชาการมีความเห็นต่าง ๆ กันเกี่ยวกับการจำแนกชั้นทางวิทยาศาสตร์ของซากที่พบ
- ↑ นักวิทยาศาสตร์บางท่านไม่คิดว่าควรจัดเป็นลิงจำพวกเอป แต่ควรจะจัดไว้ในอีกหน่วยอนุกรมวิธานที่เป็นพี่น้องกัน[118]
- ↑ หลักฐานที่ "Conroy et al., 1992"[120] จัดเข้ากับเอป ไม่ได้จัดเข้าวงศ์ลิงใหญ่
- ↑ นักวิทยาศาสตร์ผู้นำทีมที่พบซากของ "Ardi" (ทิม ไวท์) กล่าวว่า hominin สามสปีชีส์นี้คือ Sahelanthropus tchadensis, Orrorin tugenensis และ Ardipithecus kadabba (5.8 ล้านปี) เป็นสัตว์สองเท้า มีสังคมที่แตกต่างไปจากเอปทั้งที่ยังมีชีวิตอยู่และสูญพันธุ์ไปแล้ว ซึ่งรู้ได้จากขนาดของฟันที่เล็กกว่าที่ไม่ได้ใช้เพื่อเป็นอาวุธ เป็นสัตว์ที่ไม่เหมือนเอปปัจจุบัน และมีลักษณะที่เฉพาะเจาะจงเหมือนกับ hominin สกุล Australopithecus ที่ตามมาเท่านั้น ส่วนหลักฐานที่พบพร้อมกับ "Ardi" (อาร์ดี้) ซึ่งจัดอยู่ในสปีชีส์ Ardipithecus ramidus (4.4 ล้านปี) แสดงว่า อาร์ดี้เป็นสัตว์สองเท้า กินทั้งพืชและสัตว์ มีฟันเขี้ยวเล็ก มีความแตกต่างของเพศที่ลดลงเทียบกับเอป ต่างจากลิงชิมแปนซีและ Australopithecus เกินกว่าที่คาดไว้ และเป็นสัตว์อยู่ในป่าโปร่งไม่ใช่ทุ่งหญ้า (ให้สังเกตว่า ในระหว่าง A. kadabba และ A. ramidus มีช่องว่างในระหว่างที่ยังไม่พบซากดึกดำบรรพ์ของ hominin ถึงประมาณ 1.2-1.4 ล้านปี) นอกจากนั้นแล้ว สามสปีชีส์นั้นก็ยังมีตัวอย่างที่น้อยมาก จนไม่สามารถกล่าวอะไรหลาย ๆ อย่างได้อย่างชัดเจน[53]
- ↑ นักวิทยาศาสตร์ครั้งหนึ่งเคยเชื่อว่า Australopithecus africanus อาจเป็นบรรพบุรุษโดยตรงของมนุษย์ แต่หลักฐานที่เกิดขึ้นมาภายหลังคัดค้านกับสมมติฐานนี้ ในปัจจุบัน นักวิทยาศาสตร์โดยมากเชื่อว่าเป็นญาติของบรรพบุรุษมนุษย์[130]
- ↑ มีการพบท่อนกระดูกขัดแหลมในชั้นหิน Member I อายุระหว่าง 1.8 ถึง 0.5 ล้านปีก่อน ในมรดกสถานแห่งชาติ Swartkrans ประเทศแอฟริกาใต้[145] ในถ้ำที่พบกระดูกของทั้ง H. habilis[146] และ Paranthropus robustus[147] ที่หาอายุได้ใกล้ ๆ กัน
- ↑ กะโหลกศีรษะเลขที่ "NG 6" ใกล้หมู่บ้าน Ngandong ประเทศอินโดนีเซีย พบในปี ค.ศ. 1931-1933 มีอายุประมาณ 27,000-53,000 ปี[31]
- ↑ 25.0 25.1 ความสัมพันธ์ระหว่าง Denisovan กับมนุษย์ปัจจุบันและ Neanderthal ยังไม่ชัดเจน คือ งานวิจัยที่หาลำดับ mtDNA ของ Denisovan ในปี ค.ศ. 2010 เสนอว่า[171]
- มนุษย์สาย Denisovan มีบรรพบุรุษเดียวกันกับมนุษย์สาย Neanderthal และสกุล Homo ประมาณ 1 ล้านปีก่อน
- Denisovan สืบสายพันธุ์มาจากมนุษย์ที่อพยพออกมาจากแอฟริกาก่อนสายพันธุ์ของมนุษย์ปัจจุบัน และหลังจาก Homo erectus ที่มาก่อนหน้านั้น
- มีบรรพบุรุษร่วมกันกับ Neanderthal ที่แยกออกจากสายพันธุ์มนุษย์ 600,000 ปีก่อน[172]
- แยกออกจากสายพันธุ์ของ Neanderthal น่าจะในตะวันออกกลางเมื่อ 400,000 ปีก่อน[136]
- ↑ ตัวอย่างขนาดสมองอื่น ๆ ของมนุษย์ปัจจุบัน
- ขนาดเฉลี่ยเพศชายที่ 1,350 ซม3[177]
- คนพื้นเมืองออสเตรเลีย 1,199 ซม3[178]
- ชาวญี่ปุ่น 1,318 ซม3[179]
- คน Zande ในแอฟริกา 1,345 ซม3[179]
- ชาวอิตาลี 1,411 ซม3[179]
- คนพื้นเมืองสหรัฐอเมริกาเผ่า Nez Perce 1,483 ซม3[179]
- ชาวอะลิวต์ 1,518 ซม3[179]
- ↑ 27.0 27.1 งานวิจัยโดยประเมินระยะเวลาการแยกสายพันธุ์ของเหาตัวและเหาศีรษะ ซึ่งนักวิชาการใช้อนุมานถึงการใช้เสื้อผ้า ประเมินเวลาการใช้เสื้อผ้าของมนุษย์ที่ระหว่าง 83,000 - 170,000 ปีก่อน[227] หรือระหว่าง 65,000 - 149,000 ก่อน[228] แต่อาจเป็นได้ว่า การใช้เสื้อผ้าเริ่มขึ้นก่อนหน้านั้นเพราะได้พบกระดูกหมีที่มีรอยของมีคมซึ่งแสดงว่ามนุษย์ได้ใช้หนังสัตว์และน่าจะเพื่อทำให้ร่างกายอบอุ่น หลักฐานมีอายุประมาณ 300,000 ปีนี้พบในสถานโบราณคดีในเยอรมนี[229] ส่วนนักโบราณคดีได้พบเข็มเย็บเสื้อผ้าทำด้วยกระดูกและงาในประเทศรัสเซียมีอายุประมาณ 30,000 ปีก่อนคริสต์ศักราช[230] และในจีนมีอายุประมาณ 23,000-30,000 ปีก่อน[231] พบใยฝ้ายย้อมสีที่อาจใช้ทำเสื้อผ้าในถ้ำก่อนประวัติศาสตร์ในประเทศจอร์เจียมีอายุ 36,000 ปีก่อน[82]
- ↑ 28.0 28.1 มีหลักฐานว่า Neanderthal เป็นมนุษย์พวกแรกที่ตั้งใจฝังคนตาย โดยฝังในหลุมตื้น ๆ พร้อมกับเครื่องมือหินและกระดูกสัตว์ (ซึ่งตีความหมายว่าเป็นเพราะมีความเชื่อเกี่ยวกับโลกหลังความตาย)[224] แต่ก็มีนักวิชาการที่คัดค้านว่า ซากเหล่านี้อาจเกิดการฝังเพราะเหตุอื่น ๆ ที่ไม่ใช่เป็นเรื่องเกี่ยวกับเรื่องความเชื่อทางจิตวิญญาณ[225] ส่วนหลักฐาน "ที่ไม่มีใครคัดค้าน" ที่เก่าแก่ที่สุดมีอายุ 100,000 ปี เป็นซากของมนุษย์ที่แต้มสีเหลืองหรือส้มหรือน้ำตาลที่ทำจากดิน พบในถ้ำ Skhul ในประเทศอิสราเอล ซึ่งมีของต่าง ๆ ฝังรวมกับผู้ตาย[226]
- ↑ ตั้งชื่อตามชั้นหินที่พบซากดึกดำบรรพ์ ที่เก่าที่สุดจะอยู่ล่างสุด
- ↑ โรคคูรุ เป็นการติดเชื้อไวรัสนอกแบบที่ระบบประสาทส่วนกลาง พบที่ประเทศปาปัวนิวกินี เป็นโรคประสาทเสื่อมที่แก้ไขไม่ได้เกิดจากพรีออนหนึ่งที่มีในมนุษย์[235]
อ้างอิง
แก้- ↑ Heng, HH (May 2009). "The genome-centric concept: resynthesis of evolutionary theory". BioEssays. 31 (5): 512–25. doi:10.1002/bies.200800182. PMID 19334004.
- ↑ Hall, B. K.; Hallgrimsson, B., บ.ก. (2008). Strickberger's Evolution (4th ed.). Jones & Bartlett. ISBN 0-7637-0066-5.
- ↑ Panno, Joseph (2005). The Cell: Evolution of the First Organism. Facts on File. ISBN 0-8160-4946-7.
- ↑ Cracraft, J.; Donoghue, M. J., บ.ก. (2005). Assembling the tree of life. Oxford University Press. ISBN 0-19-517234-5.
- ↑ 5.0 5.1 Fleagle, J; Gilbert, C (2011). "Primate Evolution". สืบค้นเมื่อ 4 September 2014.
- ↑ "Summary - Hominidae vs Hylobatidae". Time Tree. สืบค้นเมื่อ 3 October 2014.
- ↑ 7.0 7.1 7.2 7.3 7.4 7.5 7.6 Brunet, Michel; Guy, Franck; Pilbeam, David; Mackaye, Hassane Taisso; และคณะ (2002-07-11). "A new hominid from the Upper Miocene of Chad, Central Africa". Nature. 418 (6894): 145–151. doi:10.1038/nature00879. ISSN 0028-0836. Full Article PDF (1.02 MB)
- ↑ 8.0 8.1 8.2 Henke, Winfried (2007). Henke, Winfried; Hardt, Thorolf; Tattersall, Ian (บ.ก.). Handbook of paleoanthropology: Phylogeny of hominids. Springer. pp. 1527–9. ISBN 978-3-540-32474-4.
- ↑ 9.0 9.1 9.2 Amos, Jonathan (1 October 2009). "Fossil finds extend human story". BBC News. สืบค้นเมื่อ 4 กันยายน 2014.
- ↑ 10.00 10.01 10.02 10.03 10.04 10.05 10.06 10.07 10.08 10.09 10.10 10.11 10.12 10.13 Bradley, B. J. (2008). "Reconstructing Phylogenies and Phenotypes: A Molecular View of Human Evolution". Journal of Anatomy. 212 (4): 337–353. doi:10.1111/j.1469-7580.2007.00840.x. PMC 2409108. PMID 18380860.
- ↑ 11.00 11.01 11.02 11.03 11.04 11.05 11.06 11.07 11.08 11.09 11.10 11.11 11.12 11.13 11.14 11.15 11.16 11.17 11.18 11.19 11.20 Wood, Bernard; Richmond, Brian G. (2000). "Human evolution: taxonomy and paleobiology" (PDF). Journal of Anatomy. 197 (1): 19–60. doi:10.1046/j.1469-7580.2000.19710019.x. PMC 1468107. PMID 10999270.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ 12.0 12.1 12.2 12.3 12.4 12.5 12.6 12.7 12.8 "Homo habilis". Australian Museum. 16 September 2013. สืบค้นเมื่อ 9 September 2014.
- ↑ 13.00 13.01 13.02 13.03 13.04 13.05 13.06 13.07 13.08 13.09 13.10 13.11 13.12 13.13 13.14 13.15 13.16 13.17 13.18 13.19 13.20 13.21 Potts, Richard B. (2006), "Human Evolution", Microsoft Encarta 2006
- ↑ 14.0 14.1 14.2 14.3 "Complex technology". Australian Museum. 23 November 2010. สืบค้นเมื่อ 9 September 2014.
- ↑ 15.00 15.01 15.02 15.03 15.04 15.05 15.06 15.07 15.08 15.09 Johnson, George B.; Raven, Peter H.; Singer, Susan; Losos, Jonathan (2002). "Biology/6e" (PDF). McGraw-Hill Higher Education. สืบค้นเมื่อ 23 February 2014.
- ↑ 16.0 16.1 O'Neil, Dennis (2012). "Early Transitional Humans". Anthropology Department, Palomar College. สืบค้นเมื่อ 20 กันยายน 014.[ลิงก์เสีย]
- ↑ Dennis, Megan Y.; Nuttle, Xander; Sudmant, Peter H.; Antonacci, Francesca; และคณะ (2012). "Evolution of Human-Specific Neural SRGAP2 Genes by Incomplete Segmental Duplication". Cell. 149 (4): 912–922. doi:10.1016/j.cell.2012.03.033. PMC 3365555. PMID 22559943.
- ↑ Madura, Tomas; Yamashita, Toshihide; Kubo, Tateki; Tsuji, Lyuji; และคณะ (2004). "Changes in mRNA of Slit–Robo GTPase-activating protein 2 following facial nerve transection". Molecular Brain Research. 123 (1–2): 76–80. doi:10.1016/j.molbrainres.2004.01.002.
- ↑ Wong, Kit; Ren, Xiu-Rong; Huang, Yang-Zhong; Xie, Yi; และคณะ (2001). "Signal Transduction in Neuronal Migration". Cell. 107 (2): 209–221. doi:10.1016/S0092-8674(01)00530-X. Full ArticlePDF
- ↑ Sudmant, Peter H.; Kitzman, Jacob O.; Antonacci, Francesca; Alkan, Can; และคณะ (2010-10-29). "Diversity of Human Copy Number Variation and Multicopy Genes". Science. 330 (6004): 641–646. doi:10.1126/science.1197005. ISSN 0036-8075. PMC 3020103. PMID 21030649.
- ↑ Dennis, Megan Y.; Nuttle, Xander; Sudmant, Peter H.; Antonacci, Francesca; และคณะ (2012). "Evolution of Human-Specific Neural SRGAP2 Genes by Incomplete Segmental Duplication". Cell. 149 (4): 912–922. doi:10.1016/j.cell.2012.03.033. Lay sumary from sciencenews.org เก็บถาวร 12 พฤศจิกายน 2012 ที่ เวย์แบ็กแมชชีน
- ↑ 22.0 22.1 Charrier, Cécile; Joshi, Kaumudi; Coutinho-Budd, Jaeda; Kim, Ji-Eun; และคณะ (2012). "Inhibition of SRGAP2 Function by Its Human-Specific Paralogs Induces Neoteny during Spine Maturation". Cell. 149 (4): 923–935. doi:10.1016/j.cell.2012.03.034. PMC 3357949. PMID 22559944.
- ↑ 23.0 23.1 Guerrier, Sabrice; Coutinho-Budd, Jaeda; Sassa, Takayuki; Gresset, Aurélie; และคณะ (2009). "The F-BAR Domain of srGAP2 Induces Membrane Protrusions Required for Neuronal Migration and Morphogenesis". Cell. 138 (5): 990–1004. doi:10.1016/j.cell.2009.06.047. PMC 2797480. PMID 19737524.
- ↑ 24.0 24.1 24.2 24.3 Swisher, Carl C.; Curtis, Garniss H; Lewin, Roger (2002). Java Man. Abacus. ISBN 0-349-11473-0.
- ↑ 25.0 25.1 25.2 25.3 Miller, Kenneth (7 December 2013). "Archaeologists Find Earliest Evidence of Humans Cooking With Fire". Discover Magazine.
- ↑ 26.0 26.1 26.2 26.3 26.4 {Garcia, Tristan; Féraud, Gilbert; Falguères, Christophe; de Lumley, Henry; และคณะ (2010). "Earliest human remains in Eurasia: New 40Ar/39Ar dating of the Dmanisi hominid-bearing levels, Georgia". Quaternary Geochronology. 5 (4): 443–451. doi:10.1016/j.quageo.2009.09.012.
- ↑ 27.0 27.1 27.2 Stringer, Chris (2003-06-12). "Human evolution: Out of Ethiopia". Nature. 423 (6941): 692–695. Bibcode:2003Natur.423..692S. doi:10.1038/423692a. ISSN 0028-0836. PMID 12802315. S2CID 26693109.
- ↑ 28.0 28.1 28.2 28.3 28.4 Johanson, D (May 2001). "Origins of modern humans: multiregional or out of Africa?". ActionBioscience. เก็บจากแหล่งเดิมเมื่อ 2010-11-22. สืบค้นเมื่อ 2014-09-09.
- ↑ "The first migrations out of Africa". Australian Museum. 2011-01-04. สืบค้นเมื่อ 9 September 2014.
- ↑ "The first modern humans in Southeast Asia". Australian Museum. 2010-01-04. สืบค้นเมื่อ 9 September 2014.
- ↑ 31.0 31.1 31.2 31.3 31.4 31.5 31.6 "Understanding Human" (PDF). Aug 2012. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 19 March 2015. สืบค้นเมื่อ 13 March 2015.
- ↑ 32.0 32.1 32.2 32.3 32.4 32.5 32.6 Alves, Isabel; Šrámková Hanulová, Anna; Foll, Matthieu; Excoffier, Laurent (2012-07-19). "Genomic Data Reveal a Complex Making of Humans". PLoS Genetics. 8 (7): e1002837. doi:10.1371/journal.pgen.1002837. ISSN 1553-7404. PMC 3400556. PMID 22829785. Full ArticlePDF
- ↑ Chase, P. G. (1994). "On symbols and the palaeolithic". Current Anthropology 35 (5) : 627-9.
- ↑ Knight, C (2010). Frey, U; Stormer, C; Willfuhr, KP (บ.ก.). Homo Novus - A Human Without Illusions (PDF). Berlin Heidelberg: Springer-Verlag. pp. 193–211.
- ↑ Durkheim, E. (1965). The Elementary Forms of the Religious Life. New York (NY) : Free Press.
- ↑ Chase. P. G. (1994). "On symbols and the palaeolithic". Current Anthropology 35 (5) : 627-9.
- ↑ Mellars, Paul (2006-06-20). "Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model". Proceedings of the National Academy of Sciences. 103 (25): 9381–9386. doi:10.1073/pnas.0510792103. ISSN 0027-8424. PMC 1480416. PMID 16772383.
- ↑ 38.0 38.1 38.2 38.3 38.4 38.5 Miller, Barbara D; Wood, Bernard; Balkansky, Andrew; Mercader, Julio; และคณะ (2006). "8 Modern Humans: Origins, Migrations, and Transitions". Anthropology (PDF). Boston, MA: Allyn & Bacon. pp. 223–253.
- ↑ 39.0 39.1 Diamond, Jarrad (2002). Guns, Germs and Steel. United States: Penguin. ISBN 0-393-03891-2. OCLC 35792200.
- ↑ 40.0 40.1 Dawkins (2005). "Archaic homo sapiens". The Ancestor's Tale (US Paperback). Boston: Mariner. ISBN 0-618-61916-X.
- ↑ Stanford, Craig; Allen, John S.; Anton, Susan C. (2012). "1". Biological Anthropology (2nd Edition). Prentice Hall. ISBN 0136011608.
- ↑ 42.0 42.1 Bones, Stones, and Genes - The Origin of Modern Humans (Poster) (JPG). Howard Hughes Medical Institute (2011 Holiday Lectures on Science). 2011. สืบค้นเมื่อ 4 September 2014.
- ↑ "FIGURE 25.1. Organization of the order Primates (จากหนังสือ Evolution)" (JPG). Cold Spring Harbor Laboratory Press. 2007. สืบค้นเมื่อ 4 กันยายน 2014.
- ↑ 44.0 44.1 Sayers, Ken; Raghanti, Mary Ann; Lovejoy, C. Owen (2012). "Human Evolution and the Chimpanzee Referential Doctrine". Annual Review of Anthropology. 41.
- ↑ 45.0 45.1 Ajit, Varki; Nelson, David L (2007). "Genomic Comparisons of Humans and Chimpanzees". Annu. Rev. Anthropol. 36: 191–209.
Sequence differences from the human genome were confirmed to be ∼1% in areas that can be precisely aligned, representing ∼35 million single base-pair differences. Some 45 million nucleotides of insertions and deletions unique to each lineage were also discovered, making the actual difference between the two genomes ∼4%.
- ↑ 46.0 46.1 Poznik, G. David; Henn, Brenna M.; Yee, Muh-Ching; Sliwerska, Elzbieta; และคณะ (2013-08-02). "Sequencing Y Chromosomes Resolves Discrepancy in Time to Common Ancestor of Males Versus Females". Science. 341 (6145): 562–565. doi:10.1126/science.1237619. ISSN 0036-8075. PMC 4032117. PMID 23908239.
- ↑ 47.0 47.1 Soares, Pedro; Ermini, Luca; Thomson, Noel; Mormina, Maru; และคณะ (2009). "Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock". The American Journal of Human Genetics. 84 (6): 740–759. doi:10.1016/j.ajhg.2009.05.001. PMC 2694979. PMID 19500773. University of Leeds - New 'molecular clock' aids dating of human migration history เก็บถาวร 2017-08-20 ที่ เวย์แบ็กแมชชีน
- ↑ 48.0 48.1 Cann RL, Stoneking M, Wilson AC; Stoneking; Wilson (1987). "Mitochondrial DNA and human evolution". Nature. 325 (6099): 31–6. Bibcode:1987Natur.325...31C. doi:10.1038/325031a0. PMID 3025745. เก็บจากแหล่งเดิมเมื่อ 22 November 2010. สืบค้นเมื่อ 13 March 2015.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ 49.0 49.1 49.2 Abi-Rached, Laurent; Jobin, Matthew J.; Kulkarni, Subhash; McWhinnie, Alasdair; และคณะ (2011-10-07). "The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans". Science. 334 (6052): 89–94. doi:10.1126/science.1209202. ISSN 0036-8075. PMC 3677943. PMID 21868630. archive (Aug 2011)}}-->
- ↑ 50.0 50.1 50.2 Freeman, Scott; Herron, Jon C. (2007). Evolutionary Analysis (4th ed.). Pearson Education, Inc. pp. 786–788. ISBN 978-0-13-227584-2.
- ↑ 51.0 51.1 Darwin, Charles (1859). On the Origin of Species by Means of Natural Selection (1st ed.). United Kingdom: John Murray. OCLC 352242.
- ↑ 52.0 52.1 Plummer, T (2004). "Flaked stones and old bones: Biological and cultural evolution at the dawn of technology" (PDF). Am. J. Phys. Anthropol. Suppl 39: 118–64. doi:10.1002/ajpa.20157. PMID 15605391. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 3 February 2013. สืบค้นเมื่อ 20 กันยายน 2014.
- ↑ 53.0 53.1 White, Tim (2010). "Human origins" (PDF). New Scientist - Instant Expert.
- ↑ 54.0 54.1 Plastino, Wolfango; Kaihola, L; Bartolomei, P; Bella, F (2001). "Cosmic background reduction in the radiocarbon measurement by scintillation spectrometry at the underground laboratory of Gran Sasso" (PDF). Radiocarbon. 43 (2A): 157–161.
- ↑ IUPAC, Compendium of Chemical Terminology, 2nd ed. (the "Gold Book") (1997). Online corrected version: (2006-) "radioactive dating".
- ↑ 56.0 56.1 Lovejoy, C. Owen (1988). "Evolution of Human walking" (PDF). Scientific American. 259 (5). doi:10.1038/scientificamerican1188-118.
- ↑ White, T.D.; Suwa, G.; Simpson, S.; Asfaw, B. (January 2000). "Jaws and teeth of Australopithecus afarensis from Maka, Middle Awash, Ethiopia". American Journal of Physical Anthropology. 111 (1): 45–68. PMID 10618588.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ "Australopithecus afarensis". Australian Museum. 30 November 2010. สืบค้นเมื่อ 9 September 2014.
- ↑ "The Laetoli Footprint Trail: 3D reconstruction from texture; archiving, and reverse engineering of early hominin gait (PREMOG - Supplementry Info)". Primate Evolution & Morphology Group (PREMOG), the Department of Human Anatomy and Cell Biology, the School of Biomedical Sciences, University of Liverpool. 18 May 2007. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 17 July 2007. สืบค้นเมื่อ 1 November 2007.
- ↑ Raichlen, David A.; Gordon, Adam D.; Harcourt-Smith, William E. H.; Foster, Adam D.; และคณะ (2010-03-22). "Laetoli Footprints Preserve Earliest Direct Evidence of Human-Like Bipedal Biomechanics". PLoS ONE. 5 (3): e9769. doi:10.1371/journal.pone.0009769. ISSN 1932-6203. PMC 2842428. PMID 20339543. Full ArticlePDF
- ↑ Becoming Human - an Interactive Documentary (Interactive Documentary). becominghuman.org. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (โปรแกรมคอมพิวเตอร์-ไมโครซอฟท์ วินโดวส์)เมื่อ 5 July 2014. สืบค้นเมื่อ 4 September 2014.
- ↑ "Humans As a Case Study for the Evidence of Evolution" [มนุษย์โดยเป็นกรณีศึกษาเกี่ยวกับหลักฐานของวิวัฒนาการ]. The TalkOrigins Archive. สืบค้นเมื่อ 3 October 2014.
- ↑ 63.0 63.1 White, Tim (2011). [rtmp://media.hhmi.org/vod//mp4:11Lect4_2000_jw.mp4 Bones, Stones and Genes: The Origin of Modern Humans] (MP4). Lecture 4 - Hominid Paleobiology. Howard Hughes Medical Institute (2011 Holiday Lecture Series).
- ↑ Linné, Carl von (1758). Systema naturæ. Regnum animale (10th ed.). pp. 18, 20. สืบค้นเมื่อ 19 November 2012.
- ↑ American Heritage Dictionary; Houghton Mifflin Company (October 2006). More word histories and mysteries: from aardvark to zombie. Houghton Mifflin Harcourt. pp. 99-. ISBN 978-0-618-71681-4. สืบค้นเมื่อ 10 November 2011.
- ↑ Frangsmyr, Tore; Lindroth, Sten; Eriksson, Gunnar; Broberg, Gunnar (1983). Linnaeus, the man and his work. Berkeley and Los Angeles: University of California Press. pp. 167, 170. ISBN 978-0-7112-1841-3.
- ↑ Gribbin, Mary; Gribbin, John (2008). Flower Hunters. Oxford: Oxford University Press. pp. 173–174. ISBN 978-0-19-956182-7.
- ↑ 68.0 68.1 "Homo neanderthalensis". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ 69.0 69.1 Bowler, Peter J. (2003). Evolution: The History of an Idea (3rd ed.). University of California Press. pp. 208–211, 214–216. ISBN 0-520-23693-9.
- ↑ Darwin, Charles (1981). The Descent of Man, and Selection in Relation to Sex. Princeton, N.J: Princeton University Press. ISBN 0-691-02369-7.
- ↑ "The Java Man skullcap". The TalkOrigins Archive. สืบค้นเมื่อ 3 October 2014.
- ↑ 72.0 72.1 72.2 "Prominent Hominid Fossils". The TalkOrigins Archive. สืบค้นเมื่อ 3 October 2014.
- ↑ 73.0 73.1 73.2 "Human Evolution" (PDF). MyAnthroLab. 2011. pp. 20–37. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 21 September 2015. สืบค้นเมื่อ 4 กันยายน 2014.
- ↑ 74.0 74.1 Sarich, V. M.; Wilson, A. C. (1967-12-01). "Immunological time scale for hominid evolution". Science (New York, N.Y.). 158 (3805): 1200–1203. doi:10.1126/science.158.3805.1200. ISSN 0036-8075. PMID 4964406.
- ↑ 75.0 75.1 Klages, Arthur (2011-06-21). "Sahelanthropus tchadensis: An Examination of its Hominin Affinities and Possible Phylogenetic Placement". The University of Western Ontario Journal of Anthropology. 16 (1). doi:10.5206/uwoja.v16i1.8884. ISSN 2561-7818.
- ↑ 76.0 76.1 Finlayson, Clive (2005). "Biogeography and evolution of the genus Homo". Trends in Ecology & Evolution. Elsevier. 20 (8): 457–463. doi:10.1016/j.tree.2005.05.0191. PMID 16701417.
- ↑ Stringer, CB; Andrews, P (March 1988). "Genetic and fossil evidence for the origin of modern humans". Science. 239 (4845): 1263–8. Bibcode:1988Sci...239.1263S. doi:10.1126/science.3125610. PMID 3125610.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ 78.0 78.1
Liu, H; Prugnolle, F; Manica, A; Balloux, F (August 2006). "A geographically explicit genetic model of worldwideDD human-settlement history". Am. J. Hum. Genet. 79 (2): 230–7. doi:10.1086/505436. PMC 1559480. PMID 16826514.
Currently available genetic and archaeological evidence is supportive of a recent single origin of modern humans in East Africa. However, this is where the consensus on human settlement history ends, and considerable uncertainty clouds any more detailed aspect of human colonization history.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ "Out of Africa Revisited". Science. 308 (5724): 921g–921g. 2005-05-13. doi:10.1126/science.308.5724.921g. ISSN 0036-8075.
- ↑ Wolpoff, MH; JN Spuhler; FH Smith; J Radovcic; G Pope; DW Frayer; R Eckhardt; G Clark (1988). "Modern Human Origins". Science. 241 (4867): 772–4. Bibcode:1988Sci...241..772W. doi:10.1126/science.3136545. PMID 3136545.
- ↑ 81.0 81.1 81.2 81.3 81.4 White, Tim D.; Asfaw, B.; DeGusta, D.; Gilbert, H.; และคณะ (2003). "Pleistocene Homo sapiens from Middle Awash, Ethiopia". Nature. 423 (6491): 742–747. Bibcode:2003Natur.423..742W. doi:10.1038/nature01669. PMID 12802332.
- ↑ 82.0 82.1 82.2 82.3 82.4 82.5 82.6 "Homo sapiens – modern humans". Australian Museum. 30 October 2015. สืบค้นเมื่อ 9 June 2017.
- ↑ Jorde, LB; Bamshad, M; Rogers, AR (Feb 1998). "Using mitochondrial and nuclear DNA markers to reconstruct human evolution". BioEssays. 20 (2): 126–36. doi:10.1002/(SICI)1521-1878(199802)202<126::AID-BIES5>3.0.CO;2-R. PMID 9631658.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ Cruciani, Fulvio; Trombetta, Beniamino; Massaia, Andrea; Destro-Bisol, Giovanni; และคณะ (2011). "A Revised Root for the Human Y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa". The American Journal of Human Genetics. 88 (6): 814–818. doi:10.1016/j.ajhg.2011.05.002. PMC 3113241. PMID 21601174. Full ArticlePDF (343&b)
- ↑ Mendez, Fernando L.; Krahn, Thomas; Schrack, Bonnie; Krahn, Astrid-Maria; และคณะ (2013). "An African American Paternal Lineage Adds an Extremely Ancient Root to the Human Y Chromosome Phylogenetic Tree". The American Journal of Human Genetics. 92 (3): 454–459. doi:10.1016/j.ajhg.2013.02.002. PMC 3591855. PMID 23453668. Full ArticlePDF (330 KB)
- ↑ Francalacci, Paolo; Morelli, Laura; Angius, Andrea; Berutti, Riccardo; และคณะ (2013-08-02). "Low-Pass DNA Sequencing of 1200 Sardinians Reconstructs European Y-Chromosome Phylogeny". Science. 341 (6145): 565–569. doi:10.1126/science.1237947. ISSN 0036-8075. PMC 5500864. PMID 23908240.
- ↑ 87.0 87.1 87.2 Lahr, Marta Mirazón; Petraglia, Mike; Stokes, Stephen; Field, Julie (2010). "Searching for traces of the Southern Dispersal: environmental and historical research on the evolution of human diversity in southern Asia and Australo-Melanesia". Uninversity of York.
- ↑ Macaulay, Vincent; Hill, Catherine; Achilli, Alessandro; Rengo, Chiara; และคณะ (2005-05-13). "Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes". Science. 308 (5724): 1034–1036. doi:10.1126/science.1109792. ISSN 0036-8075.
- ↑ Boyd, Robert; Silk, Joan B. (2003). How Humans Evolved. New York, New York: Norton. ISBN 0-393-97854-0.
{{cite book}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ 90.0 90.1 Brues, Alice M.; Snow, Clyde C. (1965). "Physical Anthropology". Biennial Review of Anthropology. 4: 1–39. ISBN 9780804717465.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ Kondō, Shirō (1985). Primate morphophysiology, locomotor analyses, and human bipedalism. Tokyo: University of Tokyo Press. ISBN 4-13-066093-4.
- ↑ "Your baby's journey during the 2nd stage of the labour". Birth.com. สืบค้นเมื่อ 9 กันยายน 2014.
- ↑ Sockol, Michael D.; Raichlen, David A.; Pontzer, Herman (July 24, 2007). "Chimpanzee locomotor energetics and the origin of human bipedalism". Proc. Natl. Acad. Sci. U.S.A. 104 (30): 12265–12269. Bibcode:2007PNAS..10412265S. doi:10.1073/pnas.0703267104. ISSN 0027-8424. PMC 1941460. PMID 17636134.
- ↑ "Lesson 10: Fetal Positions and Adaptations". Medical Education Division, Brookside Associates. สืบค้นเมื่อ 9 กันยายน 2014.
- ↑ Vaginal Childbirth (Birth) (ภาพยนตร์). Nucleus Medical Media. สืบค้นเมื่อ 14 October 2014.
- ↑ Strickberger, Monroe W (2005). Evolution. Jones & Bartlett Learning. pp. 475–476. ISBN 978-0-7637-3824-2. สืบค้นเมื่อ 3 October 2014.
- ↑ Wenda R. Trevathan (1 May 2011). Human Birth: An Evolutionary Perspective. Transaction Publishers. p. 20. ISBN 978-1-4128-4819-0. สืบค้นเมื่อ 12 July 2013.
- ↑ James R. Curry (2 October 2008). Children of God: Children of Earth. AuthorHouse. pp. 106–109. ISBN 978-1-4678-6390-2. สืบค้นเมื่อ 12 July 2013.
- ↑ Morris, Desmond (1967). "SEX". The Naked Ape: A Zoologist's Study of the Human Animal. ISBN 0-07-043174-4.
- ↑ Wright, Robert (1994). "Chapter 3: Men and Women". The Moral Animal (1st edition, hardcover ed.). United States: Vintage Books. ISBN 0-679-76399-6.
- ↑ Allen, L. L.; Bridges, P. S.; Evon, D. L.; Rosenberg, K. R.; และคณะ (1982). "Demography and Human Origins". American Anthropologist. 84 (4): 888–896. doi:10.1525/aa.1982.84.4.02a00140. ISSN 0002-7294.
- ↑ 102.0 102.1 Alexander, Richard D (1990). "HOW DID HUMANS EVOLVE? Reflections on the Uniquely Unique Species" (PDF). Museum Of Zoology, The University Of Michigan. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 11 April 2012. สืบค้นเมื่อ 13 March 2015.
- ↑ 103.0 103.1 103.2 103.3 103.4 103.5 103.6 Schoenemann, P. Thomas (2006). "Evolution of the Size and Functional Areas of the Human Brain" (PDF). Annu. Rev. Anthropol. 35: 379–406. doi:10.1146/annurev.anthro.35.081705.123210. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 9 June 2015. สืบค้นเมื่อ 20 กันยายน 2014.
- ↑ 104.00 104.01 104.02 104.03 104.04 104.05 104.06 104.07 104.08 104.09 104.10 "Homo neanderthalensis - The Neanderthals". Australian Museum. 25 February 2013. สืบค้นเมื่อ 3 October 2014.
- ↑ Organ, Chris; Nunn, Charles L.; Machanda, Zarin; Wrangham, Richard W. (2011-08-30). "Phylogenetic rate shifts in feeding time during the evolution of Homo". Proceedings of the National Academy of Sciences. 108 (35): 14555–14559. doi:10.1073/pnas.1107806108. ISSN 0027-8424. PMC 3167533. PMID 21873223. Full Article PDF (586 KB)
- ↑ Miller, Geoffrey (April 2007). "Ovulatory cycle effects on Tip earnings by lap dancers: economic evidence for human estrus?" (PDF). Evolution and Human Behaviour (28): 375–381.
- ↑ Ogden; และคณะ (October 27, 2004). "Mean Body Weight, Height, and Body Mass Index, United States 1960-2002" (PDF). Advance Data from Vital and Health Statistics (347).
- ↑ 108.0 108.1 Ward, C. V.; Tocheri, M. W.; Plavcan, J. M.; Brown, F. H.; และคณะ (2013). "Early Pleistocene third metacarpal from Kenya and the evolution of modern human-like hand morphology". Proceedings of the National Academy of Sciences. 111: 121. doi:10.1073/pnas.1316014110.
- ↑ Laitman, J.T.; Noden, D.M.; Van De Water, T.R. (2006). "Formation of the larynx: from homeobox". ใน Rubin, J.S.; Sataloff, R.T.; Korovin, G.S. (บ.ก.). Diagnosis & Treatment Voice Disorders genes to critical periods. San Diego: Plural. pp. 3–20. ISBN 9781597560078.
- ↑ Laitman, J.T.; Reidenberg, J.S. (2009). "The evolution of the human larynx: Nature's great experiment". ใน Fried, M.P.; Ferlito, A. (บ.ก.). The Larynx (3rd ed.). San Diego: Plural. pp. 19–38. ISBN 1597560626.
- ↑ Beard, K.C. (2008). "The oldest North American primate and mammalian biogeography during the Paleocene-Eocene Thermal Maximum" (PDF). Proceedings of the National Academy of Sciences. 105 (10): 3815. doi:10.1073/pnas.0710180105. PMC 2268774. PMID 18316721. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 2009-01-31. สืบค้นเมื่อ 13 March 2015.
- ↑ Chatterjee, Helen J; Ho, Simon YW; Barnes, Ian; Groves, Colin (2009). "Estimating the phylogeny and divergence times of primates using a supermatrix approach". BMC Evolutionary Biology. 9 (1): 259. doi:10.1186/1471-2148-9-259. ISSN 1471-2148. PMC 2774700. PMID 19860891. Full Article PDF (538 KB)
- ↑ 113.0 113.1 113.2 113.3 Begun, David R (2010). "Miocene Hominids and the Origins of the African Apes and Humans" (PDF). Annual Review of Anthropology. 39: 67–84. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 14 March 2016. สืบค้นเมื่อ 9 กันยายน 2014.
- ↑ 114.0 114.1 114.2 114.3 Hunt, Kathleen (17 March 1997). "Transitional Vertebrate Fossils FAQ - Part 2A". The TalkOrigins Archive.
- ↑ 115.0 115.1 Tyson, Peter (2008-01-07). "Meet Your Ancestors". NOVA scienceNOW (PBS).
- ↑ Zalmout, Iyad S.; Sanders, William J.; MacLatchy, Laura M.; Gunnell, Gregg F.; และคณะ (2010). "New Oligocene primate from Saudi Arabia and the divergence of apes and Old World monkeys". Nature. 466 (7304): 360–364. doi:10.1038/nature09094. ISSN 0028-0836.
- ↑ Fleagle, John G. (25 September 1998). Primate Anatomy and Evolution (second ed.). Academic Press. pp. 491–495. ISBN 978-0-12-260341-9.
- ↑ Andrews, Peter; Martin, Lawrence (1987). "Cladistic relationships of extant and fossil hominoids". Journal of Human Evolution. 16 (1): 101–118. doi:10.1016/0047-2484(87)90062-5.
- ↑ 119.0 119.1 119.2 Willoughby, Pamela R. (2005). "Palaeoanthropology and the Evolutionary Place of Humans in Nature" (PDF). International Journal of Comparative Psychology. 18 (1): 60–91.
- ↑ Haaramo, Mikko (2005-01-14). "Hominoidea - Great apes and humans". Mikko's Phylogeny Archive.
- ↑ Kunimatsu, Yutaka; Nakatsukasa, Masato; Sawada, Yoshihiro; Sakai, Tetsuya; และคณะ (2007-12-04). "A new Late Miocene great ape from Kenya and its implications for the origins of African great apes and humans". Proceedings of the National Academy of Sciences. 104 (49): 19220–19225. doi:10.1073/pnas.0706190104. ISSN 0027-8424. PMC 2148271. PMID 18024593. Full Article PDF (483 KB)
- ↑ de Bonis, L.; Bouvrain, G.; Geraads, D.; Koufost, G. (1990). "New hominid skull material from the late Miocene of Macedonia in Northern Greece". Nature. 345 (6277): 712–714. doi:10.1038/345712a0. ISSN 0028-0836.
- ↑ Stanford, Craig B. (2012). "Chimpanzees of Ardipithecus ramidus*". Anthropology. 41: 139. doi:10.1146/annurev-anthro-092611-145724.
Is Ardipithecus a hominin - that question will likely dominate the paleoanthropological debate over this fossil taxon for years to come.
- ↑ Pickford, Martin (December 2001). "Martin Pickford answers a few questions about this month's fast breaking paper in field of Geosciences". Essential Science Indicators.
- ↑ Skelton, RR; McHenry, HM (1992). "Evolutionary relationships among early hominids". Journal of Human Evolution. 23: 309–349.
- ↑ "Australopithecus afarensis". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ 127.0 127.1 Asfaw, B; White, T; Lovejoy, O; Latimer, B; และคณะ (1999). "Australopithecus garhi: a new species of early hominid from Ethiopia" (PDF). Science. 284 (5414): 629–635. doi:10.1126/science.284.5414.629. PMID 10213683. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 28 July 2018. สืบค้นเมื่อ 14 October 2014.
- ↑ "Australopithecus sediba". Australian Museum. 2011-01-20. สืบค้นเมื่อ 9 September 2014.
- ↑ "human evolution - Hominid Evolution". The Columbia Electronic Encyclopedia (6th ed.). Columbia University Press. 2012. สืบค้นเมื่อ 3 October 2014.
- ↑ "Australopithecus africanus". Australian Museum. 2011-01-20. สืบค้นเมื่อ 9 September 2014.
- ↑ 131.0 131.1 131.2 "Homo heidelbergensis". Australian Museum. 11 September 2013. สืบค้นเมื่อ 9 September 2014.
- ↑ 132.0 132.1 132.2 Hazarika, Manji (16–30 June 2007). "Homo erectus/ergaster and Out of Africa: Recent Developments in Paleoanthropology and Prehistoric Archaeology".
{{cite news}}: CS1 maint: date format (ลิงก์) - ↑ 133.0 133.1 "Homo ergaster". Australian Museum. 25 February 2013. สืบค้นเมื่อ 9 September 2014.
- ↑ 134.0 134.1 134.2 "Homo antecessor". Australian Museum. 16 September 2013. สืบค้นเมื่อ 9 September 2014.
- ↑ Wood, B (1996). "Human evolution". BioEssays. 18 (12): 945–954. doi:10.1002/bies.950181204. PMID 8976151. สืบค้นเมื่อ 9 กันยายน 2014.
- ↑ 136.0 136.1 136.2 136.3 136.4 Stringer, Chris (2012). "What makes a modern human". Nature. 485 (7396): 33–35. doi:10.1038/485033a. ISSN 0028-0836.
- ↑ Figure 1. Phylogeny of Primate Lice from Morphological and Molecular Data, in Reed, DL; Smith, VS; Hammond, SL; Rogers, AR; Clayton, DH (2004). "Genetic Analysis of Lice Supports Direct Contact between Modern and Archaic Humans". PLoS Biol. 2 (11): e340. doi:10.1371/journal.pbio.0020340. PMC 521174. PMID 15502871.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ Bryson 2004, pp. 522–543.
- ↑ van Zinderen-Bakker, E. M. (14 April 1962). "A Late-Glacial and Post-Glacial Climatic Correlation between East Africa and Europe". Nature. 194 (4824): 201–203. Bibcode:1962Natur.194..201V. doi:10.1038/194201a0.
- ↑ "The new batch - 150,000 years ago". BBC - Science & Nature - The evolution of man. เก็บจากแหล่งเดิมเมื่อ 31 July 2010. สืบค้นเมื่อ 4 September 2014.
- ↑ 141.0 141.1 141.2 "Homo rudolfensis". Australian Museum. 3 December 2009. สืบค้นเมื่อ 9 September 2014.
- ↑ Miller, J.M.A. (2000). "Craniofacial variation in Homo habilis: an analysis of the evidence for multiple species". American Journal of Physical Anthropology. 112 (1): 103–128. doi:10.1002/(SICI)1096-8644(200005)112:1<103::AID-AJPA10>3.0.CO;2-6. PMID 10766947.
- ↑ Wood, Bernard (3 April 2014). "Fifty Years After Homo habilis". Nature: 31–33.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Pollard, Elizabeth. Worlds Together, Worlds Apart. 500 Fifth Avenue, New York, N.Y. 10110. p. 11. ISBN 978-0-393-91847-2.
{{cite book}}: CS1 maint: location (ลิงก์) - ↑ "Bone Tools". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ "SK 847". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ "SK 54". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ Spoor, F; Gunz, P; Neubauer, S; Stelzer, S; Scott, N; Kwekason, A; Dean, MC (2015). "Reconstructed Homo habilis type OH 7 suggests deep-rooted species diversity in early Homo". Nature. 519 (7541): 83–86. doi:10.1038/nature14224. PMID 25739632.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Mayell, Hillary. "Killer Cats Hunted Human Ancestors". National Geographic News. สืบค้นเมื่อ 15 February 2008.
- ↑ Haeusler, Martin (Apr 2004). "Body Proportions of Homo Habilis Reviewed". Journal of Human Evolution. 46 (4): 433–465.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Tattersall, I; Schwartz, JH (2001). Extinct Humans. New York: Westview Press. p. 111.
{{cite book}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Urquhart, James (8 August 2007). "Finds test human origins theory". BBC News. สืบค้นเมื่อ 27 July 2007.
- ↑ Curnoe, D (2010). "A review of early Homo in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (Homo gautengensis sp. nov.)". HOMO - Journal of Comparative Human Biology. 61: 151–177. News: "Toothy Tree-Swinger May Be Earliest Human". Discovery News. 21 May 2010. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2 May 2012. สืบค้นเมื่อ 3 October 2014.
- ↑ Villmoare, Brian; Kimbel, William H.; Seyoum, Chalachew; Campisano, Christopher J.; และคณะ (2015-03-20). "Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia". Science. 347 (6228): 1352–1355. doi:10.1126/science.aaa1343. ISSN 0036-8075.
See also
- "Oldest known member of human family found in Ethiopia". New Scientist. 4 March 2015. สืบค้นเมื่อ 7 March 2015.,
- Ghosh, Pallab (4 March 2015). "'First human' discovered in Ethiopia". bbc.co.uk. สืบค้นเมื่อ 7 March 2015.
- ↑ "Vertebrate fossils record a faunal turnover indicative of more open and probable arid habitats than those reconstructed earlier in this region, in broad agreement with hypotheses addressing the role of environmental forcing in hominin evolution at this time."
DiMaggio, Erin N; Campisano, CJ; Rowan, J; Dupont-Nivet, G; Deino, AL และคณะ (2015). "Late Pliocene fossiliferous sedimentary record and the environmental context of early Homo from Afar, Ethiopia". Science. 347: 1355–9. doi:10.1126/science.aaa1415. PMID 25739409.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Spoor, F.; Wood, B.; Zonneveld, F. (1994-06-23). "Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion". Nature. 369 (6482): 645–648. doi:10.1038/369645a0. ISSN 0028-0836. PMID 8208290.
- ↑ Turner W (1895). "On M. Dubois' Description of Remains recently found in Java, named by him Pithecanthropus erectus: With Remarks on so-called Transitional Forms between Apes and Man". Journal of Anatomy and Physiology. 29 (Pt 3): 424–45. PMC 1328414. PMID 17232143.
- ↑ 158.0 158.1 "Homo erectus". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ F. Spoor, M. G. Leakey, P. N. Gathogo, F. H. Brown, S. C. Anton, I. McDougall, C. Kiarie, F. K. Manthi & L. N. Leakey (9 August 2007). "Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya". Nature. 448 (7154): 688–691. doi:10.1038/nature05986. PMID 17687323.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ "Fire-Altered Stone Tools". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ Wrangham, Richard (2009). Catching Fire. Basic Books.
- ↑ Fedigan, Linda Marie (1986). "The Changing Role of Women in Models of Human Evolution". Annual Review of Anthropology. 151: 25–66.
- ↑ Adrienne Zihlman; Nancy Tannerl; Lionel Tiger; Heather T. Fowler (1972). "Gathering and the Hominid Adaptation". Female Hierarchies. Beresford Book Service. pp. 220–229.
- ↑ "Hearths & Shelters". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ Gabounia L. de Lumley M. Vekua A. Lordkipanidze D. de Lumley H. (2002). "Discovery of a new hominid at Dmanisi (Transcaucasia, Georgia)". Comptes Rendus Palevol. 1 (4): 243–53. doi:10.1016/S1631-0683(02)00032-5.
- ↑ Lordkipanidze D, Vekua A, Ferring R; และคณะ (2006). "A fourth hominin skull from Dmanisi, Georgia". The anatomical record. Part A, Discoveries in molecular, cellular, and evolutionary biology. 288 (11): 1146–57. doi:10.1002/ar.a.20379. PMID 17031841.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ "Homo antecessor". Natural History Musuem. สืบค้นเมื่อ 3 October 2014.
- ↑ "Ceprano (Exhibit item)". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ Manzi, Giorgio (2012). "On the trail of the genus Homo between archaic and derived morphologies". Journal of Anthropological Sciences (90): 1–18. doi:10.4436/jass.90018. ISSN 1827-4765. Full Article PDF (2.19 MB)
- ↑ Schuster, Angela M.H. "New Skull from Eritrea". Archaeology Magazine Archive. สืบค้นเมื่อ 2024-03-13.
- ↑ Sample, Ian (24 March 2010). "New species of human ancestor found in Siberia". The Guardian. สืบค้นเมื่อ 20 กันยายน 2014.
- ↑ 172.0 172.1 Reich, David; Green, Richard E.; Kircher, Martin; Krause, Johannes; และคณะ (2010-12-23). "Genetic history of an archaic hominin group from Denisova Cave in Siberia". Nature. 468 (7327): 1053–1060. doi:10.1038/nature09710. ISSN 1476-4687. PMC 4306417. PMID 21179161.
- ↑ 173.0 173.1 O'Neil, Dennis (2012). "Homo heidelbergensis". Anthropology Department, Palomar College. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 9 March 2016. สืบค้นเมื่อ 20 กันยายน 2014.
- ↑ Carretero, José-Miguel; Rodríguez, Laura; García-González, Rebeca; Arsuaga, Juan-Luis; และคณะ (2012). "Stature estimation from complete long bones in the Middle Pleistocene humans from the Sima de los Huesos, Sierra de Atapuerca (Spain)". Journal of Human Evolution. 62 (2): 242–255. doi:10.1016/j.jhevol.2011.11.004. ScienceDaily Summary - June 6, 2012
- ↑ Burger, Lee (November 2007). "Our Story: Human Ancestor Fossils". The Naked Scientists.
- ↑ Rightmire, G. Philip (2005). "The Lake Ndutu cranium and early Homo Sapiens in Africa" (PDF). American Journal of Physical Anthropology. 61 (2): 245–254. doi:10.1002/ajpa.1330610214. PMID 6410925. สืบค้นเมื่อ 14 October 2014.
- ↑ "If Modern Humans Are So Smart, Why Are Our Brains Shrinking?". Discover Magazine. 2011-01-20.
- ↑ Klekamp, J; Riedel, A; Harper, C; Kretschmann (February 1987). "A quantitative study of Australian aboriginal and Caucasian brains". J Anat. 150: 191–210. PMC 1261675. สืบค้นเมื่อ 26 กันยายน 2014.
- ↑ 179.0 179.1 179.2 179.3 179.4 Smith, Courtland L; Beals, Kenneth L (1990). "Cultural Correlates with Cranial Capacity". American Anthropologist. 92 (1): 193–200. doi:10.1525/aa.1990.92.1.02a00150.
- ↑ "Neanderthal Brain Size at Birth Sheds Light on Human Evolution". National Geographic. 9 September 2008. สืบค้นเมื่อ 19 September 2009.
- ↑ Viegas, Jennifer (6 May 2010). "Neanderthals, humans interbred, DNA proves". Discovery News. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 18 August 2010. สืบค้นเมื่อ 17 August 2010.
- ↑ Hebsgaard, Martin B.; Wiuf, Carsten; Gilbert, M. Thomas P.; Glenner, Henrik; Willerslev, Eske (2007). "Evaluating Neanderthal Genetics and Phylogeny". Journal of Molecular Evolution. 64 (1): 50–60. doi:10.1007/s00239-006-0017-y. ISSN 0022-2844.
- ↑ Gutiérrez, Gabriel; Sánchez, Diego; Marín, Antonio (2002-08-01). "A Reanalysis of the Ancient Mitochondrial DNA Sequences Recovered from Neandertal Bones" (PDF). Molecular Biology and Evolution. 19 (8): 1359–1366. doi:10.1093/oxfordjournals.molbev.a004197. ISSN 1537-1719.
- ↑ "Science & Nature Wildfacts—Neanderthal". BBC. สืบค้นเมื่อ 21 June 2009.
- ↑ Mellars, Paul; French, Jennifer C. (2011-07-29). "Tenfold Population Increase in Western Europe at the Neandertal–to–Modern Human Transition". Science. 333 (6042): 623–627. doi:10.1126/science.1206930. ISSN 0036-8075.
- ↑ 186.0 186.1 186.2 "Homo floresiensis". Australian Museum. 11 September 2013. สืบค้นเมื่อ 3 October 2014.
- ↑ Martin RD, Maclarnon AM, Phillips JL, Dobyns WB (2006). "Flores hominid: new species or microcephalic dwarf?". The anatomical record. 288 (11): 1123–45. doi:10.1002/ar.a.20389. PMID 17031806.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ Muehlenbein, Michael (2010). Human Evolutionary Biology. Cambridge University Press. pp. 192–213.
- ↑ Frisancho, AR (1993). Human Adaptation and Accommodation. University of Michigan Press. pp. 175–301. ISBN 0472095110.
- ↑ Mayel, Hillary (24 February 2004). "Three High-Altitude Peoples, Three Adaptations to Thin Air". National Geographic News. National Geographic Society. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2 July 2013. สืบค้นเมื่อ 13 March 2015.
- ↑ 191.0 191.1 191.2 191.3 Richter, Daniel; Grün, Rainer; Joannes-Boyau, Renaud; Steele, Teresa E.; และคณะ (2017-06-08). "The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age". Nature. 546 (7657): 293–296. doi:10.1038/nature22335. ISSN 0028-0836. Abstract
- ↑ 192.0 192.1 Hublin, Jean-Jacques; Ben-Ncer, Abdelouahed; Bailey, Shara E.; Freidline, Sarah E.; และคณะ (2017-06-08). "New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens". Nature. 546 (7657): 289–292. doi:10.1038/nature22336. ISSN 0028-0836.
- ↑ 193.0 193.1 193.2 "Oldest Homo sapiens fossil claim rewrites our species' history". Nature News. 7 June 2017.
- ↑ 194.0 194.1 194.2 194.3 194.4 "Oldest Fossils of Homo Sapiens Found in Morocco, Altering History of Our Species". เดอะนิวยอร์กไทมส์. 7 June 2017.
- ↑ 195.0 195.1 195.2 "Earliest known human fossils found in Morocco". Nature Middle East. 7 June 2017.
- ↑ 196.0 196.1 "New fossil finds from Morocco do more than push back the origins of our species by 100,000 years". Science Daily. 7 June 2017.
- ↑ 197.0 197.1 197.2 "World's oldest Homo sapiens fossils found in Morocco". Science Magazine. 7 June 2017.
- ↑ 198.0 198.1 198.2 198.3 "Oldest Homo sapiens fossils discovered in Morocco". Washington Post. 7 June 2017.
- ↑ Jabr, Ferris (18 July 2012). "Does Thinking Really Hard Burn More Calories?". Scientific American.
- ↑ Richards, M.P. (December 2002). "A brief review of the archaeological evidence for Palaeolithic and Neolithic subsistence (full article)". European Journal of Clinical Nutrition. 56 Supplement 1, March 2002 (12): 1270–1278. doi:10.1038/sj.ejcn.1601646. PMID 12494313.
{{cite journal}}: CS1 maint: date and year (ลิงก์) Full Article PDF (132 KB) - ↑ Gibbons, Ann (May 29, 1998). "Solving the Brain's Energy Crisis". Science. 280 (5368): 1345–1347. doi:10.1126/science.280.5368.1345. ISSN 0036-8075. PMID 9634409. S2CID 28464775.
- ↑ "Humans are apes - 'Great Apes'". Australian Museum. 1 March 2013. สืบค้นเมื่อ 9 September 2014.
- ↑ 203.0 203.1 "Early Stone Age Tools". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ 204.0 204.1 204.2 "Middle Stone Age Tools". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ Skinner; และคณะ (2007). "New ESR Dates for a New Bone-Bearing Layer at Pradayrol, Lot, France". Abstracts of the PaleoAnthropology Society 2007 Meetings. PaleoAnthropology: A1–A35.
- ↑ Shea, J. J. (2003). "Neandertals [sic], competition and the origin of modern human behaviour in the Levant". Evolutionary Anthropology. 12: 173–187.
- ↑ 207.0 207.1 Langer, William L., บ.ก. (1972). An Encyclopedia of World History (5th ed.). Boston, MA: Houghton Mifflin Company. p. 9. ISBN 0-395-13592-3.
- ↑ Tryon, Christopher A; Faith, Tyler (2013). "Variability in the Middle Stone Age of Eastern Africa". Current Anthropology. 54 (8): 234–254. doi:10.1086/673752.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) - ↑ Wenke, Robert J; Olszewski, Deborah (2012). "4 The Origins of Homo sapiens sapiens". Patterns in Prehistory : Humankind's First Three Million Years (PDF). คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 1 September 2015. สืบค้นเมื่อ 3 October 2014.
{{cite book}}: CS1 maint: multiple names: authors list (ลิงก์) - ↑ 210.0 210.1 "New dates for old cave art in Spain". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ Than, Ker (14 June 2012). "World's Oldest Cave Art Found—Made By Neanderthals?". National Geographic Society. สืบค้นเมื่อ 30 December 2012.
- ↑ Bailey, Martin (2013-01-31). "Ice Age Lion Man is world's earliest figurative sculpture". The Art Newspaper. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2014-08-10. สืบค้นเมื่อ 2013-02-01.
- ↑ "Earliest music instruments found". BBC News. 25 May 2012.
- ↑ "12 The Origin and Dispersal of Modern Human". Paleoanthropology/Fossil Hominins (PDF). 2012. pp. 284–310. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 19 March 2015. สืบค้นเมื่อ 13 March 2015.
- ↑ 215.0 215.1 Henshilwood, Christopher S.; และคณะ (2011). "A 100,000-Year-Old Ochre-Processing Workshop at Blombos Cave, South Africa". Science. 334: 219–222.
- ↑ 216.0 216.1 Henshilwood, Christopher S.; d'Errico, Francesco; Yates, Royden; Jacobs, Zenobia; และคณะ (2002-02-15). "Emergence of Modern Human Behavior: Middle Stone Age Engravings from South Africa". Science. 295 (5558): 1278–1280. doi:10.1126/science.1067575. ISSN 0036-8075.
- ↑ Marean, Curtis W.; Bar-Matthews, Miryam; Bernatchez, Jocelyn; Fisher, Erich; และคณะ (2007). "Early human use of marine resources and pigment in South Africa during the Middle Pleistocene". Nature. 449 (7164): 905–908. doi:10.1038/nature06204. ISSN 0028-0836.
- ↑ http://www.baanjomyut.com/library/human_evolution/
- ↑ http://www.manager.co.th/Science/ViewNews.aspx?NewsID=9510000128144[ลิงก์เสีย]
- ↑ Diamond, Jared (1999). Guns, Germs, and Steel: The Fate of Human Societies. W. W. Norton. p. 39. ISBN 978-0-393-31755-8.
- ↑ Bar-Yosef, Ofer (2002). "The Upper Paleolithic Revolution" (PDF). Annual Review of Anthropology. 31: 363–393.[ลิงก์เสีย]
- ↑
d'Errico, Francesco; Stringer, Chris B (12 April 2011). "Evolution, revolution or saltation scenario for the emergence of modern cultures?" (PDF). Phil. Trans. R. Soc. B. 366 (1567): 1060–1069. doi:10.1098/rstb.2010.0340.
{{cite journal}}: CS1 maint: date and year (ลิงก์) - ↑ Nowell, April (2010). "Defining Behavioral Modernity in the Context of Neandertal and Anatomically Modern Human Populations". Annual Review of Anthropology. 39: 437–452. doi:10.1146/annurev.anthro.012809.105113.
- ↑ "The World's Largest Neanderthal Finding Site". Culturenet Iaccessdate=2010-08-07.
- ↑ "Evolving in their graves: early burials hold clues to human origins - research of burial rituals of Neanderthals". Science News. The Free Library. 15 December 2001. สืบค้นเมื่อ 3 October 2014.
- ↑ Lieberman, Philip (1993). Uniquely Human. Harvard University Press. pp. 163–164. ISBN 9780674921832.
- ↑ Toups, M. A.; Kitchen, A.; Light, J. E.; Reed, D. L. (2011-01-01). "Origin of Clothing Lice Indicates Early Clothing Use by Anatomically Modern Humans in Africa". Molecular Biology and Evolution. 28 (1): 29–32. doi:10.1093/molbev/msq234. ISSN 0737-4038. PMC 3002236. PMID 20823373. Full ArticlePDF (237 KB)
- ↑ Kittler, Ralf; Kayser, Manfred; Stoneking, Mark (2003-08-19). "Molecular evolution of Pediculus humanus and the origin of clothing". Current biology: CB. 13 (16): 1414–1417. doi:10.1016/s0960-9822(03)00507-4. ISSN 0960-9822. PMID 12932325.
- ↑ Hamer, Ashley (2024-03-10). "When did humans start wearing clothes?". livescience.com. เก็บจากแหล่งเดิมเมื่อ 2024-03-12. สืบค้นเมื่อ 2024-03-13.
- ↑ Hoffecker, J; Scott, J (21 March 2002). "Excavations In Eastern Europe Reveal Ancient Human Lifestyles". University of Colorado at Boulder News Archive. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 19 July 2011. สืบค้นเมื่อ 13 March 2015.
- ↑ "Bone and Ivory Needles (Exhibit Item)". Smithsonian Institution. สืบค้นเมื่อ 4 October 2014.[ลิงก์เสีย]
- ↑ William H. Calvin (1 September 2003). "A Brief History of the Mind". Oxford University Press. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2018-06-26. สืบค้นเมื่อ 10 September 2009.
- ↑ Henshilwood, Christopher S; Marean, Curtis W (December 2003). "The Origin of Modern Human Behavior: Critique of the Models and Their Test Implications". Current Anthropology. 44 (5): 627–651. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 19 July 2022. สืบค้นเมื่อ 4 กันยายน 2014.
{{cite journal}}: CS1 maint: bot: original URL status unknown (ลิงก์) - ↑ Mellars, Paul (2006-06-20). "Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model". Proceedings of the National Academy of Sciences. 103 (25): 9381–9386. doi:10.1073/pnas.0510792103. ISSN 0027-8424. PMC 1480416. PMID 16772383. Full ArticlePDF
- ↑
Wadsworth, JD และคณะ (March 2008). "Kuru prions and sporadic Creutzfeldt-Jakob disease prions have equivalent transmission properties in transgenic and wild-type mice". Proc. Natl. Acad. Sci. USA. 105 (10): 3885–90. doi:10.1073/pnas.0800190105. PMC 2268835. PMID 18316717.
{{cite journal}}: CS1 maint: uses authors parameter (ลิงก์) Full ArticlePDF - ↑ Medical Research Council (UK) (November 21, 2009). "Brain Disease 'Resistance Gene' evolves in Papua New Guinea community; could offer insights Into CJD". Science Daily (online). Science News. เก็บจากแหล่งเดิมเมื่อ 22 November 2010. สืบค้นเมื่อ 22 November 2009.
- ↑ Mead, Simon; Whitfield, Jerome; Poulter, Mark; Shah, Paresh; และคณะ (2009-11-19). "A Novel Protective Prion Protein Variant that Colocalizes with Kuru Exposure". New England Journal of Medicine. Massachusetts Medical Society. 361 (21): 2056–2065. doi:10.1056/nejmoa0809716. ISSN 0028-4793.
- ↑ Byars, Sean G.; Ewbank, Douglas; Govindaraju, Diddahally R.; Stearns, Stephen C. (2010-01-26). "Natural selection in a contemporary human population". Proceedings of the National Academy of Sciences. 107 (suppl_1): 1787–1792. doi:10.1073/pnas.0906199106. ISSN 0027-8424. PMC 2868295. PMID 19858476. Full Article PDF (1.03 MB)
- ↑ Cochran, G; Harpending, H (2009). The 10,000 Year Explosion. New York: Basic Books. ISBN 0-465-00221-8.
- ↑ "Understanding Human" (PDF). Aug 2012. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 19 March 2015. สืบค้นเมื่อ 4 กันยายน 2014.
- ↑ Sahi, Timo (2001). "Genetics and epidemiology of adult-type hypolactasia with emphasis on the situation in Europe". Näringsforskning. 45 (1): 161–162. doi:10.3402/fnr.v45i0.1799. ISSN 0027-7878.
- ↑ 242.0 242.1 Kretchmer, N (1972). "Lactose and lactase". Sci. Am. 227 (4): 71–8. PMID 4672311.
- ↑ "Introduction to Cladistics". University of California, Berkeley. สืบค้นเมื่อ 2014-01-06.
- ↑ Merriam-Webster Collegiate Dictionary, 11th Edition. Springfield, Massachusetts, USA: Merriam-Webster, Inc. 2003.
- ↑ Strait, David S.; Grine, Frederick E. (December 2004). "Inferring hominoid and early hominid phylogeny using craniodental characters: the role of fossil taxa" (PDF). Journal of Human Evolution. 47 (6): 399–452. doi:10.1016/j.jhevol.2004.08.008.
- ↑ Organ, Chris; Nunn, Charles L.; Machanda, Zarin; Wrangham, Richard W. (2011-08-30). "Phylogenetic rate shifts in feeding time during the evolution of Homo". Proceedings of the National Academy of Sciences. 108 (35): 14555–14559. doi:10.1073/pnas.1107806108. ISSN 0027-8424. PMC 3167533. PMID 21873223.
{kind=link}
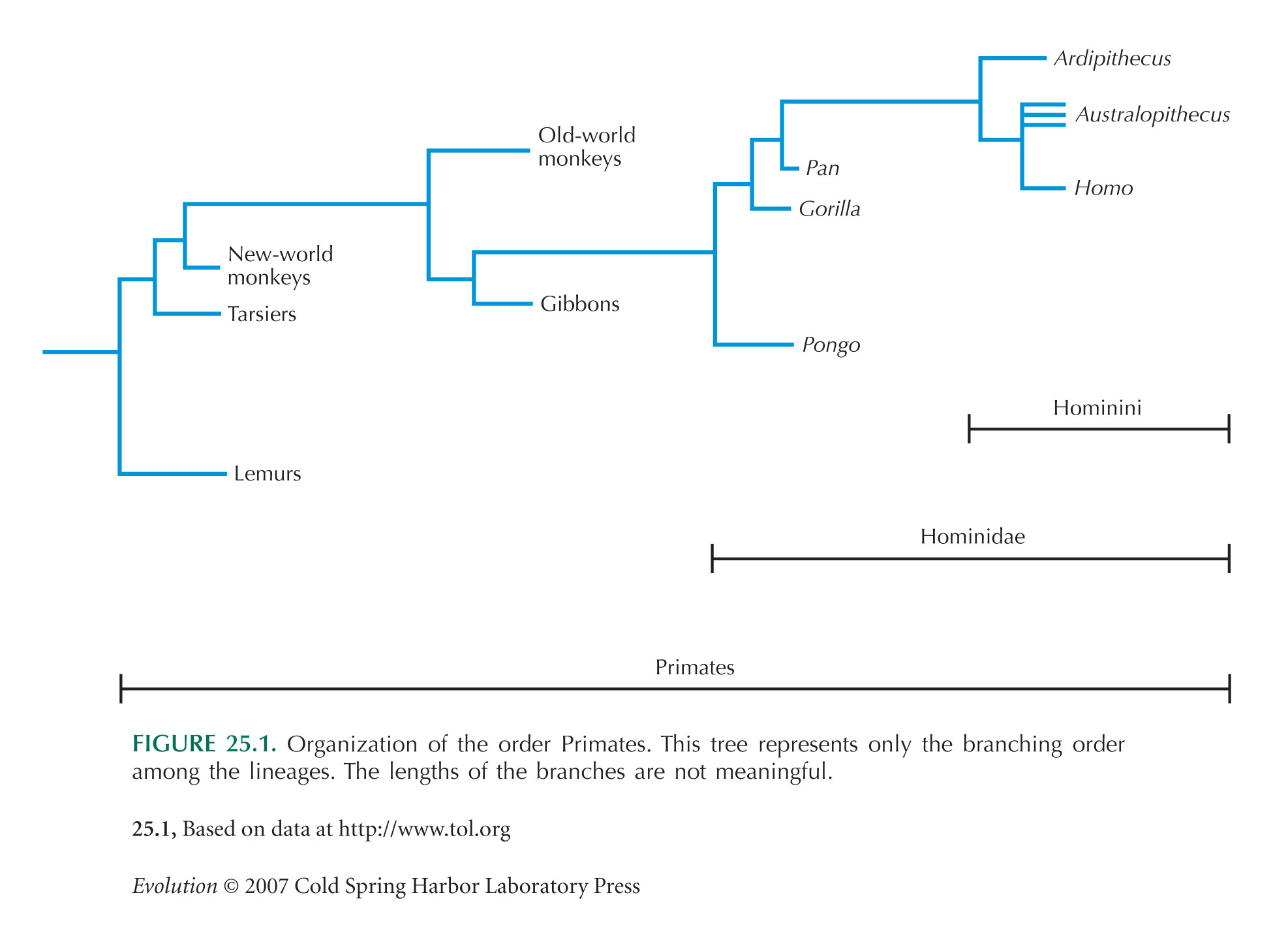{kind=link}
แหล่งข้อมูลอื่น
แก้แหล่งข้อมูลแบ่งเป็นสองส่วนเพราะว่าหลักฐานเริ่มตั้งแต่ปี ค.ศ. 2000 มักจะมีข้อมูลทางพันธุศาสตร์รวมเข้ามาด้วย
ตั้งแต่ปี ค.ศ. 2000
แก้- Enard, Wolfgang; Przeworski, Molly; Fisher, Simon E.; Lai, Cecilia S. L.; และคณะ (22 August 2002). "Molecular evolution of FOXP2, a gene involved in speech and language". Nature. 418 (6900): 869-72 [870]. doi:10.1038/nature01025. PMID 12192408.
- Flinn, Mark V.; Geary, David C.; Ward, Carol V. (2005). "Ecological dominance, social competition, and coalitionary arms races". Evolution and Human Behavior. 26 (1): 10–46. doi:10.1016/j.evolhumbehav.2004.08.005. Full Article PDF (345 KB)
- Gibbons, Ann (2007). The First Human: The Race to Discover our Earliest Ancestor. Anchor Books. ISBN 978-1-4000-7696-3.
- Hartwig, Walter Carl (2008-09-18). The Primate Fossil Record. Cambridge University Press. ISBN 978-0-521-08141-2.
- Heizmann, Elmar P J, Begun, David R (2001). "The oldest Eurasian hominoid". Journal of Human Evolution. 41 (5): 463–81. doi:10.1006/jhev.2001.0495. PMID 11681862.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์) - Johanson, Donald; Wong, Kate (2009). Lucy's Legacy: The Quest for Human Origins. Three Rivers Press. ISBN 978-0-307-39640-2.
- Morwood, Mike; van Oosterzee, Penny (2007). A New Human: The Startling Discovery and Strange Story of the 'Hobbits' of Flores, Indonesia. Smithsonian Books. ISBN 978-0-06-089908-0.
- Oppenheimer, Stephen (2003). Out of Eden: The Peopling of the World. Constable. ISBN 1-84119-697-5.
- Ovchinnikov; Götherström, Anders; Romanova, Galina P.; Kharitonov, Vitaliy M.; Lidén, Kerstin; Goodwin, William; และคณะ (2000). "Molecular analysis of Neanderthal DNA from the Northern Caucasus". Nature. 404 (6777): 490–3. doi:10.1038/35006625. PMID 10761915.
- Roberts, Alice (2009). The Incredible Human Journey: The Story of how we Colonised the Planet. Bloomsbury. ISBN 978-0-7475-9839-8.
- Stringer, Chris (2011). The Origin of Our Species. Allen Lane. ISBN 978-1-84614-140-9.
- Tattersall, Ian (2008). The Fossil Trail: How We Know What We Think We Know About Human Evolution. Oxford University Press. ISBN 978-0195367669.
- Wade, Nicholas (2006). Before the Dawn: Recovering the Lost History of our Ancestors. Penguin Press. ISBN 978-0-7156-3658-9.
- "Oldest Human Skeleton and the Missing Link". National Geographic. 1 October 2009. สืบค้นเมื่อ 4 กันยายน 2014.
ก่อนปี ค.ศ. 2000
แก้- Alexander, R. D. (1990). "How did humans evolve? Reflections on the uniquely unique species" (PDF). University of Michigan Museum of Zoology Special Publication. University of Michigan Museum of Zoology (1): 1–38. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 11 April 2012. สืบค้นเมื่อ 13 March 2015.
- Hill, Andrew; Ward, Steven (1988). "Origin of the hominidae: The record of African large hominoid evolution between 14 my and 4 my". Yearbook of Physical Anthropology. 31 (59): 49–83. doi:10.1002/ajpa.1330310505.
- Ijdo, J. W; Baldini, A; Ward, D. C; Reeders, S. T; Wells, R. A; Baldini; Ward; Reeders; Wells (October 1991). "Origin of human chromosome 2: An ancestral telomere-telomere fusion" (PDF). Genetics. 88 (20): 9051–9055. Bibcode:1991PNAS...88.9051I. doi:10.1073/pnas.88.20.9051. PMC 52649. PMID 1924367.
{{cite journal}}: CS1 maint: multiple names: authors list (ลิงก์)—two ancestral ape chromosomes fused to give rise to human chromosome 2 - Jones, Steve; Martin, Robert D.; Pilbeam, David R, บ.ก. (1994). The Cambridge Encyclopedia of Human evolution. Cambridge University Press. ISBN 978-0-521-46786-5. (Note: this book contains very useful, information dense chapters on primate evolution in general, and human evolution in particular, including fossil history).
- Leakey, Richard; Lewin, Roger (1992). Origins Reconsidered: In Search of What Makes us Human. Little, Brown and Company. ISBN 0-316-90298-5.
- Lewin, Roger (1987). Bones of Contention: Controversies in the Search for Human Origins. Penguin Books. ISBN 0-14-022638-9.
- Shreeve, James (1996). The Neanderthal Enigma: Solving the Mystery of Modern Human Origins. Viking. ISBN 0-670-86638-5.
- Stringer, Chris; McKie, Robin (1996). African Exodus: The Origins of Modern Humanity. Jonathan Cape. ISBN 0-224-03771-4.
- Walker, Allan; Shipman, Pat (1996). The Wisdom of the Bones: In Search of Human Origins. Weidenfeld and Nicholson. ISBN 0-297-81670-5.
- Weiss, M.L., & Mann, A. E (1985). 'Human Biology and Behaviour: An anthropological perspective (4th ed.). Boston: Little Brown. ISBN 978-0-673-39013-4.
{{cite book}}: CS1 maint: multiple names: authors list (ลิงก์) (Note: this book contains very accessible descriptions of human and non-human primates, their evolution, and fossil history).
แหล่งข้อมูลทางเว็บไซต์
แก้- "Human Evolution". Smithsonian Institution Human Origins Program. เป็นเว็บไซต์สื่อประสมภาษาอังกฤษที่อ่านง่ายและมีข้อมูลทันสมัย
- "Human Evolution". Australian Musuem. เป็นเว็บไซต์ภาษาอังกฤษมีข้อมูลทันสมัย
- "Bones, Stones, and Genes - The Origin of Modern Humans". Howard Hughes Medical Institute (2011 Holiday Lectures on Science). 2011. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 17 April 2012. สืบค้นเมื่อ 13 March 2015. มีวิดีโอเล็กเช่อร์พื้นฐาน (มุ่งกลุ่มเยาวชน) ภาษาอังกฤษเกี่ยวกับการศึกษาวิวัฒนาการมนุษย์ของนักวิชาการชื่อดังรวมทั้งทิม ไวท์, Sarah Tishkoff และ John Shea
- รายการ The Evolution of Man ของ BBC
- "Illustrations from Evolution (textbook)" [รูปจากตำรา Evolution (วิวัฒนาการ) ]. Evolution.
- Becoming Human: Paleoanthropology, Evolution and Human Origins, presented by Arizona State University's Institute of Human Origins
- "Human Evolution Timeline". ArchaeologyInfo.com. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 18 June 2013. สืบค้นเมื่อ 3 October 2014.